
Nature子刊:CAR-T细胞疗法目前的局限性和潜在策略
来源:本站原创 2021-04-17 06:11
2021年4月17日讯/生物谷BIOON/---嵌合抗原受体(CAR)T细胞(CAR-T)疗法是革命性的,因为它已经产生了非常有效和持久的临床反应。CAR是一种工程化的合成受体,其功能是重新引导淋巴细胞(最常见的是T细胞)识别和消除表达特定靶抗原的细胞。CAR与细胞表面上表达的靶抗原的结合不依赖于MHC受体,从而导致强有力的T细胞活化和强大的抗肿瘤反应。抗C
2021年4月17日讯/生物谷BIOON/---嵌合抗原受体(CAR)T细胞(CAR-T)疗法是革命性的,因为它已经产生了非常有效和持久的临床反应。CAR是一种工程化的合成受体,其功能是重新引导淋巴细胞(最常见的是T细胞)识别和消除表达特定靶抗原的细胞。CAR与细胞表面上表达的靶抗原的结合不依赖于MHC受体,从而导致强有力的T细胞活化和强大的抗肿瘤反应。抗CD19 CAR-T细胞疗法治疗B细胞恶性肿瘤的空前成功导致它在2017年被美国食品药品管理局(FDA)批准上市。然而,CAR-T细胞疗法仍有重大的局限性必须解决,包括危及生命的CAR-T细胞相关毒性,对实体瘤的疗效有限,B细胞恶性肿瘤中的抑制和抵抗性,抗原逃逸,有限的持久性,较差的迁移和肿瘤浸润,以及免疫抑制性微环境。此外,技术人员必须适应这个不断增长和发展的领域的需要,为此需要制定教育计划来培训他们。

人们已经提出许多方法,包括将CAR-T细胞疗法与其他抗癌疗法相结合,或采用创新的CAR工程策略,以提高抗肿瘤疗效,扩大临床疗效,并限制毒性。在这篇综述文章中,美国威斯康星大学麦迪逊分校的Robert C. Sterner和梅奥诊所的Rosalie M. Sterner讨论了CAR-T细胞工程的最新创新,以提高对血液恶性肿瘤和实体瘤的临床疗效,以及克服当前存在的包括抗原逃逸、CAR-T细胞迁移、肿瘤浸润、免疫抑制性微环境和CAR-T细胞相关毒性在内的局限性的策略(图1)。

图1.CAR-T细胞疗法的局限性。图片来自Blood Cancer Journal, 2021, doi:10.1038/s41408-021-00459-7。
CAR结构
CAR是模块化的合成受体,主要由四个部分组成:(1)一个细胞外靶抗原结合结构域(extracellular target antigen-binding domain);(2)一个铰链区(hinge region),(3)一个跨膜结构域(transmembrane domain);(4)一个或多个细胞内信号转导结构域(intracellular signaling domain)。本文中,这两名作者将讨论目前CAR设计的基本原则。
(1)抗原结合结构域。抗原结合结构域是CAR中赋予靶抗原特异性的部分。历史上,抗原结合结构域来源于单克隆抗体的可变重链(VH)和轻链(VL),通一个柔性的连接物连接VH和VL可形成单链可变片段(scFv)。通常而言,存在于CAR中的scFv靶向细胞外表面癌症抗原,导致不依赖于主要组织相容性复合体(MHC)的T细胞活化,尽管使用MHC依赖性的模拟T细胞受体(TCR)的 CAR识别细胞内肿瘤相关抗原已经被描述。除了简单地识别和结合靶表位之外,scFv的几个特点影响CAR功能。比如,VH和VL链之间的相互作用模式,以及互补性决定区(CDR)的相对位置影响CAR对其靶表位的亲和力和特异性。亲和力是抗原结合结构域的一个特别重要的参数,因为它从根本上决定了CAR的功能。为了识别肿瘤细胞表面上的抗原、诱导CAR信号转导并激活T细胞,CAR的抗原结合亲和力必须足够高,但不能高到足以导致CAR-T细胞遭受激活诱导的细胞死亡,并引发毒性。虽然亲和力肯定是使问题进一步复杂化的最重要因素之一,但已有研究表明,即使具有相似亲和力的scFv也可以差异性地影响CAR-T细胞功能。因此,为了优化CAR与靶抗原的结合,必须考虑其他因素,如表位位置,靶抗原密度,以及避免使用与配体无关的补强信号(tonic signaling)相关的scFv。
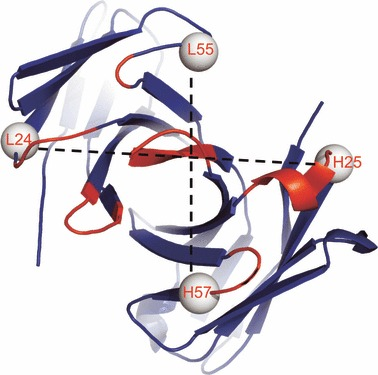
抗原结合位点空间示意图,图片来自FEBS Journal, 2011, doi:10.1111/j.1742-4658.2011.08207.x。
(2)铰链区。铰链或间隔区被定义为从跨膜结构域延伸结合单元的细胞外结构区域。铰链区的功能是提供灵活性,以克服空间阻碍,并对CAR的长度有贡献,以便允许抗原结合结构域接触到靶表位。重要的是,所选择的铰链区似乎影响CAR的功能,因为铰链区的长度和组成的差异可以影响灵活性、CAR表达、信号转导、表位识别、激活输出的强度和表位识别。除了这些影响之外,已经人提出,铰链区长度对于提供足够的细胞间距离以形成免疫突触是至关重要的。原则上,“最佳”铰链区长度取决于目标细胞上的靶表位位置和空间阻碍水平:长的铰链区提供增加的灵活性,并允许更有效地接触到膜近端表位或复杂的糖基化抗原,而短的铰链区更成功地结合膜远端表位。然而,在实践中,合适的铰链区长度往往是经验性确定的,必须为每对特定的抗原结合结构域定制。文献中有许多例子,比如短间隔CAR[CD19和癌胚抗原(CEA)]和长间隔CAR[粘蛋白1(MUC1),受体酪氨酸激酶样孤儿受体1(ROR1)的膜近端表位]。最常用的铰链区来自CD8,CD28,IgG1或IgG4的氨基酸序列。然而,源自IgG的铰链区可以导致CAR-T细胞耗竭,从而降低在体内的持久性,因为它们可以与Fcγ受体相互作用。这些影响可以通过选择不同的间隔区或通过基于功能或结构的考虑对间隔区进行进一步的改造来避免。
(3)跨膜结构域。在CAR的所有组件中,跨膜结构域可能是最少被表征的区域。跨膜结构域的主要功能是将CAR锚定在T细胞膜上,尽管有证据表明,跨膜结构域也可以与CAR-T细胞功能相关。更具体地说,有研究表明,CAR跨膜结构域影响CAR的表达水平和稳定性,活跃于信号转导或突触形成过程中,并与内源性信号分子二聚化。大多数跨膜结构域来自天然蛋白,包括CD3ζ、CD4、CD8α或CD28。不同跨膜结构域对CAR功能的影响没有得到很好的研究,这是因为跨膜结构域经常根据细胞外间隔区或细胞内信号转导结构域的要求而改变。
值得注意的是,CD3ζ跨膜结构域可能促进CAR介导的T细胞激活,因为CD3ζ跨膜结构域介导CAR二聚化和整入内源性TCR中。与具有CD28跨膜结构域的CAR相比,CD3ζ跨膜结构域的这些有益影响是以降低CAR稳定性为代价的。跨膜结构域和铰链区似乎也影响CAR-T细胞的细胞因子产生和激活诱导的细胞死亡(AICD),这是因为与具有来自CD28的跨膜结构域和铰链区的CAR相比,具有CD8α跨膜结构域和铰链区的CAR-T细胞释放的TNF和IFNγ的数量减少,对AICD的敏感性降低。总的来说,有研究表明,将近端胞内结构域与相应的跨膜结构域连接在一起,可能最能促进CAR-T细胞信号的正确传递,而使用常用的CD8α或CD28跨膜结构域可能会增强CAR的表达和稳定性。
(4)细胞内信号转导结构域。可以说,在CAR工程中最受关注的是了解CAR共刺激的效果,目标是生成具有最佳胞内结构域的CAR构建体。在20世纪90年代后期设计出的第一代CAR包含CD3ζ或FcRγ信号转导结构域。绝大多数CAR依赖于通过CD3ζ衍生性免疫受体酪氨酸激活基序来激活CAR-T细胞。然而,有效的T细胞反应是不能够仅通过这些激活基序的信号转导产生。这些第一代CAR的持久性在体外并不强劲。这些发现得到了显示出有限或没有疗效的临床研究的呼应。
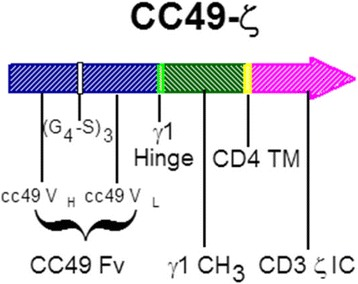
CC49-ζ 载体图,图片来自J Immunother Cancer, 2017, doi:10.1186/s40425-017-0222-9。
使用B细胞恶性肿瘤的早期体内模型证实了共刺激在靶向CD-19的CAR-T细胞持久性方面的重要性。通过添加共刺激结构域,重复抗原暴露后的IL-2产生和细胞增殖得到了改善。随着对共刺激对持久性CAR-T细胞疗法的重要性的理解,人们构建出具有一个与CD3ζ细胞内信号转导结构域串联的共刺激结构域的第二代CAR。两个最常见的获得FDA批准的共刺激结构域CD28和4-1BB(CD137)都与较高的患者反应率相关。这两种共刺激结构域的功能和代谢谱不同:具有CD28结构域的CAR-T细胞分化为效应记忆T细胞,主要使用有氧糖酵解,而具有4-1BB结构域的CAR-T细胞分化为中枢记忆T细胞,并显示出增加的线粒体生物生成和氧化代谢。
临床上,第二代CAR-T细胞在一些血液恶性肿瘤中产生了强烈的治疗反应,包括慢性淋巴细胞白血病、B细胞急性淋巴细胞白血病、弥漫性大B细胞淋巴瘤和多发性骨髓瘤。目前正在研究第二代CAR-T细胞在实体瘤中的疗效,包括胶质母细胞瘤、晚期肉瘤、肝转移瘤以及间皮瘤、卵巢癌和胰腺癌。几种替代性共刺激结构域,如诱导性T细胞共刺激物(ICOS)、CD27、MYD88、CD40和OX40(CD134)已显示出临床前的疗效,尽管临床研究仍有待进行。据推测,仅通过一个结构域的共刺激产生不完全的激活,这导致了第三代CAR的产生。第三代CAR整入两个与CD3ζ串联的共刺激结构域。第三代CAR的临床前研究产生了好坏参半的结果。具体来说,整入CD28和4-1BB信号的CAR在淋巴瘤中导致更强的细胞因子产生,与第二代CAR相比,它们在肺转移瘤中显示出更好的体内抗肿瘤反应。在白血病和胰腺癌模型中,第三代CAR没有显示出体内治疗优势,并且在各自的模型中未能优于第二代CAR。
CAR-T细胞治疗的局限性
(1)抗原逃逸。CAR-T细胞疗法最具挑战性的局限性之一是肿瘤对单抗原靶向CAR(即靶向单抗原的CAR)构建体产生抵抗性。虽然最初的单抗原靶向CAR-T细胞可以提供高反应率,但在接受这些CAR-T细胞治疗的患者中,相当一部分患者的恶性细胞表现出部分或完全丧失靶抗原表达。这种现象称为抗原逃逸。例如,尽管70%~90%的复发和/或难治性ALL患者对CD19靶向CAR-T细胞治疗表现出持久的反应,但最近的随访数据表明,存在一种常见的疾病抵抗性机制,包括30%~70%的在治疗后有复发疾病的患者出现CD19抗原下调/丧失。类似地,在正在接受BCMA靶向CAR-T细胞治疗的多发性骨髓瘤患者中,BCMA表达的下调或丧失已被观察到。类似的抗原逃逸抵抗性模式已在实体瘤中观察到。比如,靶向胶质母细胞瘤中IL13Ra2的CAR-T细胞治疗病例报告表明,肿瘤复发时IL13Ra2表达减少。
为了降低CAR-T细胞治疗血液恶性肿瘤和实体瘤的复发率,现在许多策略都依赖于靶向多种抗原。这些策略都采用了双CAR构建体或串联CAR的方法,串联CAR是指含有两个scFv的单个CAR构建体,以便同时靶向多种肿瘤靶抗原。从临床上来看,这两种策略都可能导致长期持久的缓解率,目前已有多项针对CD19和CD20或CD19和CD22的临床试验。令人振奋的是,使用双靶向CAR-T细胞(CD19/CD22或CD19/BCMA)的临床试验的初步结果显示了有希望的结果。更具体地说,CD19/CD22 CAR-T细胞疗法的初步临床试验结果在ALL和弥漫性大B细胞淋巴瘤成人患者中表现出了良好的疗效。此外,BCMA/CD19靶向CAR-T细胞治疗多发性骨髓瘤的初步结果表明,BCMA/CD19靶向CAR具有较高的疗效和良好的安全性。在实体瘤中,已经在临床前模型中测试了几种串联CAR,包括靶向胶质母细胞瘤中的HER2和IL13Ra2的串联CAR以及靶向乳腺癌中的HER2和MUC1的串联CAR。在这两种情况下,相比于单靶向疗法,双重靶向导致了优越的抗肿瘤反应。在胶质母细胞瘤研究中,与其他两种双靶向疗法相比,靶向HER2和IL13Ra2的串联CAR导致抗肿瘤活性改善,抗原逃逸减少。这项研究说明了优化选择靶抗原的重要性,这不仅可以提高抗肿瘤反应,还可以减少抗原逃逸,防止复发。
(2)非肿瘤组织的在靶效应(On-target off-tumor effects,以下简称脱靶效应)。靶向实体瘤抗原的挑战之一是,实体瘤抗原通常也在正常组织中以不同水平表达。因此,抗原选择在CAR设计中至关重要,不仅能保证治疗效果,还能限制脱靶毒性。克服靶向也存在于正常组织中的实体肿瘤抗原的一个潜在途径是靶向仅限于肿瘤的翻译后修饰,如实体瘤过表达的截短O -聚糖,比如Tn(GalNAca1-O-Ser/Thr)和sialyl-Tn(STn)(NeuAca2–6-GalNAca1-O-Ser/Thr)。四个主要的CAR-T细胞靶标已被研究,包括TAG7228,B7-H3,MUC1和MUC16。虽然第一代靶向结直肠癌中TAG72的CAR-T细胞没有产生抗肿瘤反应,但目前正在研究第二代TAG72-CAR-T细胞的新版本和其他肿瘤限制性翻译后修饰。为了扩大CAR-T细胞疗法在血液恶性肿瘤和实体瘤中的临床应用,进一步开发创新策略以减少抗原逃逸和选择能够诱导足够的抗肿瘤疗效的抗原,同时将毒副作用降到最低将是必要的。
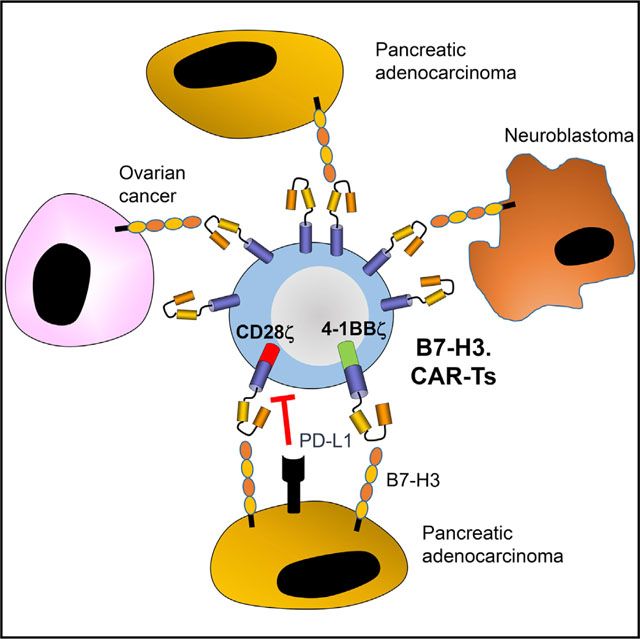
靶向B7-H3的CAR-T细胞,图片来自Cancer Cell, 2019, doi:10.1016/j.ccell.2019.01.002。
(3)CAR-T细胞的迁移和肿瘤浸润。与血液恶性肿瘤相比,实体瘤CAR-T细胞治疗受限于CAR-T细胞向实体瘤迁移和浸润的能力,因为免疫抑制性的肿瘤微环境和物理性的肿瘤屏障,如肿瘤基质,限制了CAR-T细胞的渗透和迁移。改善这些局限性的一个策略是利用除全身给送以外的给送途径,这是因为局部给送(1)消除了CAR-T细胞向疾病部位迁移的需要,(2)限制了脱靶毒性,毕竟CAR-T细胞的在靶活性是针对肿瘤细胞的,从而最大限度地减少它们与正常组织的相互作用。临床前模型已经证明,在乳腺癌脑转移和胶质母细胞瘤中,靶向HER2的CAR-T细胞和靶向IL13Ra2的CAR-T细胞的静脉注射具有卓越的治疗效果。这些研究已经导致了三项正在进行的临床试验,这三项临床试验分别针对胶质母细胞瘤(NCT02208362,NCT03389230)和复发性脑或柔脑膜转移(NCT03696030)进行CAR-T细胞的静脉注射。同样,临床前模型显示,通过胸膜内注射CAR-T细胞治疗恶性胸膜间皮瘤具有优越性,这已经导致了一项正在进行的1期临床试验(NCT02414269)。虽然局部注射似乎有较好的疗效,但理论上这种方法仅限于单个肿瘤病灶/寡转移性疾病(oligometastatic disease)。
最近开发的一种策略似乎可以显著改善CAR-T细胞的迁移,涉及在CAR-T细胞表面上表达趋化因子受体,以匹配和响应肿瘤源性趋化因子。例如,最近的研究表明,经基因修饰后表达CXCR2的整合素αvβ6-CAR-T细胞或过度表达CXCR1或CXCR2的CAR-T细胞都增强了迁移和显著提高抗肿瘤疗效。肿瘤基质等物理屏障也限制了CAR-T细胞疗法,这是因为这些物理屏障阻止了对肿瘤的渗透。肿瘤基质主要由细胞外基质组成,而在细胞外基质中,硫酸肝素蛋白聚糖(HSPG)是CAR-T细胞必须降解从而进入肿瘤的主要成分。经过基因改造后表达降解HSPG的乙酰肝素酶(heparanase)的CAR-T细胞显示出增强的肿瘤浸润和抗肿瘤活性。同样,靶向成纤维细胞活化蛋白(FAP)的CAR-T细胞在动物模型中通过减少肿瘤成纤维细胞表现出增加的细胞毒性功能。在未来,有必要开发创新的递送策略和方法,以提高肿瘤渗透,以便将治疗效果扩大到复杂的实体瘤和转移瘤。
(4)免疫抑制性微环境。在肿瘤微环境中,许多驱动免疫抑制的细胞类型可以浸润实体瘤,包括骨髓源性抑制细胞(MDSC)、肿瘤相关巨噬细胞(TAM)和调节性T细胞(Treg)。这些肿瘤浸润细胞驱动肿瘤促进细胞因子、趋化因子和生长因子的产生。此外,免疫检查点途径,如PD-1或CTLA-4,可用于降低抗肿瘤免疫。对CAR-T细胞治疗无反应或反应较弱的主要原因之一是T细胞扩增较差和T细胞的短期持续存在。据推测,这种T细胞功能衰竭的产生由共同抑制途径触发。因此,CAR-T细胞和免疫检查点阻断的联合免疫治疗被认为是下一个免疫治疗的前沿,因为它提供了强大免疫反应所必需的两个要素:(1)浸润肿瘤的CAR-T细胞;(2)PD-1/PD-L1阻断,可确保维持T细胞的持久性和功能。在血液恶性肿瘤中,在宾州儿童医院的一项单中心研究中,对14名事先接受大量治疗的B-ALL儿童进行PD-1阻断和CD19 CAR-T细胞联合治疗,改善了CAR-T细胞的持久性和实现了更好的临床结果。在实体瘤中,目前有许多研究旨在评估这类联合治疗的反应率。在一项耐人寻味的研究中,11名间皮瘤患者在接受环磷酰胺预处理后,再接受单剂量的间皮素靶向CAR-T细胞和至少三剂抗PD-1药物,结果实现72%的反应率和两名患者的完全代谢反应。结合其他形式的免疫治疗策略可能仍然是必要的,以便对抗存在于肿瘤微环境中的抑制信号。
最近的研究工作集中在对不友好的肿瘤微环境中的免疫抑制因子(比如TGF β介导的抑制信号)有抵抗力的CAR基因改造上。另一个有趣的策略涉及对CAR-T细胞进行基因改造,以刺激性细胞因子的形式提供免疫刺激信号,增加T细胞的生存、增殖、抗肿瘤活性,并重新平衡肿瘤微环境。许多研究已经探究了许多细胞因子来创建这些“装甲CAR(armored CAR)”。关注促炎细胞因子的表达而不是关注抑制信号的研究依赖于IL-12分泌、IL-15表达,以及将免疫抑制性细胞因子(如IL-4)信号重新引向促炎细胞因子。
虽然免疫检查点阻断-CAR-T细胞联合疗法很可能是一种新的免疫治疗选择,但也要认识到,即使这种组合可能仍然不足以诱导T细胞的浸润和效应功能。因此,将CAR-T细胞疗法和免疫检查点阻断与其他免疫疗法/策略相结合的额外研究可能是实现T细胞在复杂血液恶性肿瘤或实体瘤中的浸润和效应功能所必需的。
(5)CAR-T细胞相关毒性。虽然CAR-T细胞疗法已经是一种革命性的癌症治疗工具,但较高的毒副作用率与一些死亡阻止了CAR-T细胞疗法成为一线治疗。可能决定细胞因子释放综合征(CRS)、嗜血细胞综合征(HLH)/巨噬细胞活化综合征(MAS)和/或免疫效应细胞相关神经毒性综合征(ICANS)发生率和严重程度的关键因素是CAR的设计、特定靶点和肿瘤类型。到目前为止,CAR-T细胞疗法的毒性已经在接受获得FDA批准的第一个CAR-T细胞疗法---靶向CD19的CAR-T细胞---的患者中得到了最广泛的描述。即使在反应率最高的临床试验中,患者也会发生严重的危及生命的事件。具体来说,在急性淋巴细胞白血病/淋巴瘤(ALL/LBL)患者接受CAR-T细胞治疗的情况下,几乎所有的患者都有至少一些不太严重的毒性表现,而23%~46%的患者表现出严重的超生理细胞因子的产生和大量的体内T细胞增殖。这些毒性水平的全身细胞因子释放和部分患者出现的严重的免疫细胞交叉激活导致以下毒性:(1)CRS,它与超生理细胞因子的产生和大量体内T细胞增殖有关;(2)HLH/MAS,它被定义为一种严重的高炎症综合征,其特征是CRS、血清铁蛋白升高和噬血细胞增多、肾功能衰竭、肝酶、脾肿大、肺水肿和/或自然杀伤(NK)细胞活性缺失;(3)ICANS,其特征是脑脊液细胞因子水平升高和血脑屏障破坏。
从机制上讲,CRS是由于给送的CAR-T细胞被广泛激活,导致大量细胞因子的释放。轻度CRS的临床表现是发热,伴有疲乏、腹泻、头痛、皮疹、关节痛和肌痛,在更严重的情况下,患者可能会出现低血压、心脏功能障碍、循环衰竭、呼吸衰竭、肾功能衰竭、多器官系统衰竭,并有可能进展到死亡。总的来说,77%~93%的接受CAR-T细胞治疗的白血病患者和37%~93%的接受CAR-T细胞治疗的淋巴瘤患者有任何一种级别的CRS,而用tisagenlecleucel治疗复发/难治性B-ALL的患者中46%的人和用axicabtagene ciloleucel和tisagenlecleucel治疗弥漫性大B细胞淋巴瘤的患者中13%~18%的人表现出≥3级CRS。在病理生理学上,CRS被认为主要是由IL-6介导的,因此,CRS管理依赖于使用托珠单抗和皮质类固醇来阻断IL-6受体。即使使用FDA批准用来治疗重度CRS的托珠单抗,重度CRS和死亡仍然发生。
有趣的是,CAR-T细胞治疗后的HLH/MAS对IL-6抑制产生抵抗性,可能需要化疗。虽然由于与高级别CRS重叠,CAR-T细胞治疗后的HLH/MAS发生率尚不清楚,但在大约1%的接受CAR-T细胞治疗的患者中已有报道。在神经毒性的情况下,潜在的病理生理学和机制尚不完全清楚。ICANS的临床表现范围从精神错乱、头痛、注意力障碍、寻字困难、局灶性神经功能障碍或脑病到危及生命的脑水肿、短暂性昏迷或癫痫发作。CAR-T细胞治疗后的神经毒性相对较为常见,在接受治疗的白血病患者和淋巴瘤患者中,分别有高达67%和62%的人发生神经毒性。对神经毒性的管理集中在皮质类固醇,因为IL-6抑制剂通常对CAR-T细胞治疗相关的神经毒性无效。到目前为止,仍然没有批准的疗法来预防上述毒性,这就使得优化CAR设计和采用其他策略来减少CAR诱导的毒性至关重要。下面,这些作者回顾了在CAR设计中的经验教训以减少毒性,以及采取的额外策略,以缓解CAR-T细胞疗法中的毒性。
针对CAR-T细胞的局限性采取的应对策略
为了达到有效的治疗反应,CAR-T细胞的抗原结合结构域必须结合它的靶表位,并达到最低阈值水平,以诱导CAR-T细胞激活和细胞因子分泌。但与此同时,也存在一定的激活阈值水平,当超过该阈值水平时,会产生毒性水平的细胞因子和免疫系统激活。换句话说,CAR-T细胞必须保持在其治疗窗口内才能发挥临床疗效,因为超过治疗窗口将导致毒性。从基因改造的角度来看,CAR-T细胞的激活程度和激活动力学受到几个因素的影响,包括但不限于恶性细胞表面上表达的肿瘤抗原的水平,肿瘤负荷,抗原结合结构域对其靶表位的亲和力,以及CAR的共刺激结构域。因此,在优化治疗效果和限制毒性方面,仔细考虑CAR的模块化结构的几个组成部分是必要的。
(1)改变CAR结构。降低毒性的一个途径是通过改变CAR-T细胞的抗原结合结构域的亲和力。降低抗原结合结构域的亲和力预计将导致对肿瘤细胞表面上更高的抗原密度的要求增加,以实现高水平的激活。因此,预计下降的抗原亲和力将规避对具有相对较低抗原量的健康组织的靶向。探究这一原理的研究表明,与具有低纳摩尔/亚纳摩尔亲和力的抗原结合结构域相比,具有微摩尔亲和力的抗原结合结构域对具有较高水平的靶抗原表达的肿瘤的选择性要高得多。
人们也有可能通过通过修饰活化的CAR-T细胞的铰链区和跨膜区来调节细胞因子分泌。例如,在靶向CD19的CAR中,修饰CD8-α衍生的铰链区和跨膜区氨基酸序列可导致细胞因子释放水平降低,CAR-T细胞增殖减少。优化铰链区和跨膜区可能是一个有用的方法来降低毒性,因为在1期临床试验中,这些在铰链区和跨膜区发生修饰的CAR导致54.5%的B细胞淋巴瘤患者(6/11名患者)完全缓解,重要的是,没有级别>1的CRS或ICANS事件发生。
共刺激结构域在CAR设计中提供了另一个可修饰的区域,该区域可以根据肿瘤类型、肿瘤负荷、抗原密度、靶抗原-抗原结合结构域对以及潜在的毒副作用进行定制。具体来说,4-1BB结构域导致较低的毒性风险,较高的T细胞耐久性,以及较低的T细胞增殖峰值水平,而CD28共刺激结构域与CAR-T细胞活性有关,该活性的起始和随后的衰竭更为迅速。因此,产生较小毒性的4-1BB共刺激结构域在高疾病负担和/或高抗原密度的肿瘤的情况下可能特别有用,而CD28共刺激结构域可能是在总表面抗原密度较低和/或具有低亲和力抗原结合结构域的CAR的情况下达到所需的T细胞活化阈值所需要的。
(2)降低免疫原性。宿主免疫系统对CAR构建体的识别可能促进细胞因子相关的毒性,因此,利用人类或人源化抗体片段代替鼠源CAR来降低CAR的免疫原性可能是有利的。此外,铰链区和/或跨膜结构域可以被修改,以降低CAR的免疫原性,而且有趣的是,CAR-T细胞的持久性得到改善。
(3)修改CAR转导的T细胞,降低神经毒性。一个令人兴奋的最近开发的防止CAR-T细胞细胞因子毒性的途径是基于对CAR转导的T细胞进行修饰。细胞因子和髓样细胞似乎在CAR-T细胞诱导的神经毒性中起着重要作用,因为有报道显示,在3级或更高的神经毒性患者中CD14+细胞显著增加,而且一项关键的针对大B细胞淋巴瘤的CAR-T细胞临床试验表明,在与3级或更高的神经毒性产生相关的血清生物标志物中,GM-CSF升高与神经毒性最显著相关。最近的临床前研究表明,在用lenzilumab抑制活化巨噬细胞和单核细胞的细胞因子GM-CSF后,神经毒性和CRS减少,CAR-T细胞活性增加。GM-CSF突变失活在CAR转导的T细胞中似乎也有类似的效果。
因此,这些研究结果表明,GM-CSF中和有助于减少神经毒性和减少CRS。此外,以髓样细胞特异性的方式剔除酪氨酸羟化酶或使用甲酪氨酸抑制这种酶,导致儿茶酚胺和细胞因子水平下降。临床前证据还表明,IL-1受体拮抗剂减少了用靶向CD19的CAR-T细胞治疗的白血病/淋巴瘤小鼠模型中的一种神经炎症形式。
(4)CAR“关闭开关”。缓解CAR-T细胞毒性的另一个潜在途径是通过实施“关闭开关”或自杀基因策略。这样的策略将有助于在不良事件发生时通过二级诱导剂处理选择性减少经过基因改造的细胞。利用这些概念的几种方法已被开发。比如,独立表达全长CD20或CD20模拟表位或经过基因改造后表达全长CD20或CD20模拟表位的CAR构建体促进通过利妥昔单抗处理剔除CAR-T细胞。然而,这种方法的一个局限性是抗体介导的CAR-T细胞剔除的相对缓慢的发生,这可能会限制这种方法在严重的急性细胞因子介导的毒性期间需要立即逆转这种毒性的患者中的疗效。这促使人们开发出更快的开关,如诱导性cas9。在一项临床试验中,诱导性cas9在30分钟内消除了90%以上经过基因改造的T细胞。
其他策略依赖于基于蛋白酶的小分子辅助关闭CAR(SMASh-CAR),这也被称为关闭开关CAR(SWIFF-CAR)。自杀策略或其他类似方法的最大局限性是,尽管它们对确保安全性很有吸引力,但它们的使用会突然停止对快速进展的疾病的治疗。这一限制已经成为一种强大的动机,促使人们制定策略来确保安全性,而自杀基因激活则是最后的手段。一种具有令人兴奋的潜力的方法涉及使用酪氨酸激酶抑制剂达沙替尼,其功能是通过抑制近端TCR信号激酶来抑制T细胞的激活。在临床前模型中,达沙替尼快速且可逆地阻止CAR-T细胞的激活,并且在CAR-T细胞输注后早期给予达沙替尼的结果是显著降低小鼠因致命性CRS出现的死亡率。因此,这种方法似乎提供了对CAR-T细胞功能的暂时性抑制,并且可以允许在毒性消退后挽救CAR-T细胞疗法。未来,开发暂时抑制CAR-T细胞功能并在毒性消退后允许CAR-T细胞疗法挽救的额外创新方法将是CAR-T细胞治疗迈向血液恶性肿瘤和实体瘤一线治疗的必要条件。
结论
CAR是一种模块化的合成受体,主要由四个部分组成:一个细胞外靶抗原结合结构域、一个铰链区、一个跨膜结构域和一个或多个细胞内信号转导结构域。CAR-T细胞已经彻底改变了某些血液恶性肿瘤的治疗。然而,仍然存在障碍。对技术人员进行培训使之满足这个复杂和不断发展的领域的需求是具有挑战性的,需要开发创新的课程。抗原选择对CAR-T细胞功能至关重要。由于CAR-T细胞的选择性压力,肿瘤细胞可以下调抗原表达。即使有适当的抗原靶向,脱靶效应也会发生,并引起相关的毒性。在实体瘤中,让CAR-T细胞迁移到肿瘤并浸润肿瘤是一大挑战。恶性肿瘤的免疫抑制性微环境会加剧这一障碍。有效的治疗也会导致CAR-T细胞相关毒性(比如CRS和神经毒性)的风险。然而,虽然存在挑战,但新的策略和潜在的解决方案仍在不断发展,并且可能为未来更有效和更安全的治疗提供一条道路。(生物谷 Bioon.com)
参考资料:
Robert C. Sterner et al. CAR-T cell therapy: current limitations and potential strategies. Blood Cancer Journal, 2021, doi:10.1038/s41408-021-00459-7.
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
