
2021年4月CRISPR/Cas研究进展
来源:本站原创 2021-04-30 14:45
2021年4月30日讯/生物谷BIOON/---基因组编辑技术CRISPR/Cas9被《科学》杂志列为2013年年度十大科技进展之一,受到人们的高度重视。2020年10月,德国马克斯-普朗克病原学研究所的Emmanuelle Charpentier博士以及美国加州大学伯克利分校的Jennifer A. Doudna博士因在CRISPR-Cas9基因编辑方面做
2021年4月30日讯/生物谷BIOON/---基因组编辑技术CRISPR/Cas9被《科学》杂志列为2013年年度十大科技进展之一,受到人们的高度重视。2020年10月,德国马克斯-普朗克病原学研究所的Emmanuelle Charpentier博士以及美国加州大学伯克利分校的Jennifer A. Doudna博士因在CRISPR-Cas9基因编辑方面做了的贡献荣获2020年诺贝尔化学奖。
CRISPR是规律间隔性成簇短回文重复序列的简称,Cas是CRISPR相关蛋白的简称。CRISPR/Cas最初是在细菌体内发现的,是细菌用来识别和摧毁抗噬菌体和其他病原体入侵的防御系统。
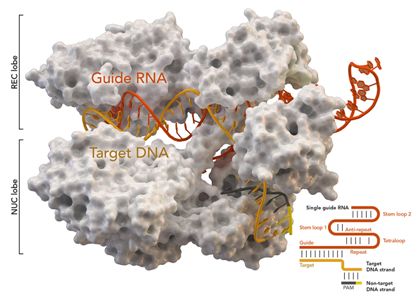
图片来自Thomas Splettstoesser (Wikipedia, CC BY-SA 4.0)。
2018年11月26日,中国科学家贺建奎声称世界上首批经过基因编辑的婴儿---一对双胞胎女性婴儿---在11月出生。他利用一种强大的基因编辑工具CRISPR-Cas9对这对双胞胎的一个基因进行修改,使得她们出生后就能够天然地抵抗HIV感染。这也是世界首例免疫艾滋病基因编辑婴儿。这条消息瞬间在国内外网站上迅速发酵,引发千层浪。有部分科学家支持贺建奎的研究,但是更多的是质疑,甚至是谴责。
即将过去的4月份,有哪些重大的CRISPR/Cas研究或发现呢?小编梳理了一下这个月生物谷报道的CRISPR/Cas研究方面的新闻,供大家阅读。
1.Nature:揭示体细胞基因组编辑的发展机遇和挑战
doi:10.1038/s41586-021-03191-1
遗传因素导致了大多数类型的人类疾病,包括遗传性疾病、传染性疾病和恶性疾病。因此,生物医学科学的一个长期目标是开发一种手段来修改患者体内的基因组,以校正致病突变,使入侵病原体的基因组失效,使免疫细胞攻击肿瘤,并使无数其他治疗机会得以实现。在某些情况下,基因添加可以具有治疗价值,而且基因疗法正在经历越来越多的成功。然而,在许多其他情况下,必须对患者的基因组进行编辑才能达到治疗效果。基因组编辑广泛地包括不同的技术,可以在不同的环境下做出许多不同的基因组改变,这个话题已经成为近期的全面综述的主题。基因组编辑中的几个概念是SCGE联盟(Somatic Cell Gene Editing Consortium, 体细胞基因编辑联盟)的目标和战略的核心。
在过去的几十年里,技术的稳步发展使得用户可编程的基因组编辑已被引入,测试,改进和实施。这些技术包括同源重组、锌指核酸酶(ZFN)、归巢核酸内切酶(meganuclease)和转录激活因子样效应物核酸酶(TALEN)。最近,源自细菌免疫途径的工程化分子复合物称为CRISPR-Cas系统,已经彻底改变了基因组编辑,部分原因是它们的靶序列可以简单地用易于设计的向导RNA进行编程。尽管有了这些有希望的进展,但在治疗性基因组编辑的转化潜力可以充分实现之前,挑战仍然存在。
在一篇新的观点(Perspective)类型文章中,作者概述了SCGE联盟的目标和战略,该联盟是由美国国家卫生研究院(NIH)成立的,旨在加快制定应对这些挑战的解决方案。NIH在6年内拨款约1.9亿美元支持SCGE联盟,该联盟如今包括来自38家研究机构的72名主要研究人员,他们正在开展45个不同但又相互结合的项目。
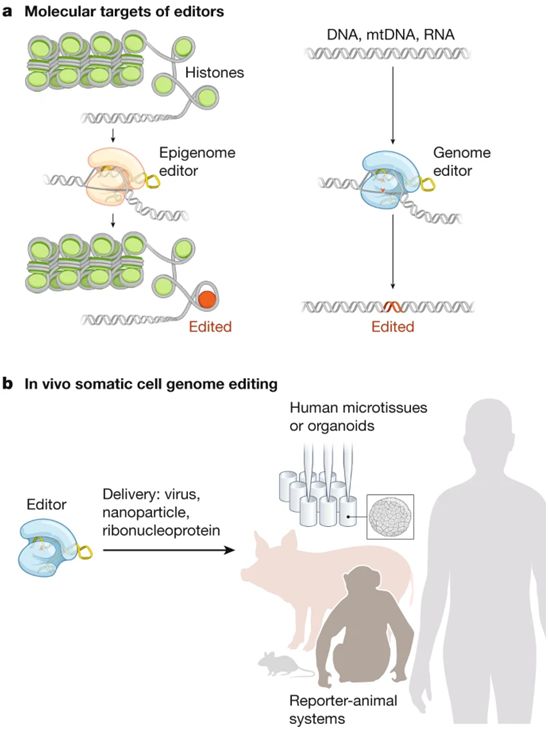
用于在体内编辑细胞基因组的工具。图片来自Nature, 2021, doi:10.1038/s41586-021-03191-1。
SCGE联盟的总体目标是加速基因组编辑技术向各种组织和疾病的转化。该领域的主要挑战之一是使用共同的指标和标准对各种技术进行比较。例如,一种视网膜递送系统可能会在感兴趣的基因处产生在靶插入或缺失(on-target indel),但不清楚这种相同的递送系统是否可以校正肺部中的不同基因。SCGE项目中交织着能够混合和匹配各种技术和读数的发展路径。在一个例子中,SCGE项目前三年开发的所有新的递送技术将首先在小动物(例如,小鼠)中进行测试,然后---如果成功的话---在猪和非人类灵长类动物中进行测试。由此产生的第三方数据将与更大的研究界和公众共享。SCGE联盟的一个关键价值是透明性,这使得其他人能够获得其研究产出,并利用其结果和产品为他们自己的疾病重点项目提供信息和加快研发进度。除了数据,这些作者旨在提供一系列工具、试剂、方法和最佳实践,这些工具、试剂、方法和最佳实践将被整合到SCGE治疗性基因组编辑工具包(简称SCGE工具包)中。通过这些活动和可交付成果,SCGE联盟力求减少开发新疗法所需的时间和成本,以便产生持久的影响。
2.Cell: 新型CRISPR转录组学编辑“机器”有助于重塑转录组记忆
doi:10.1016/j.cell.2021.03.025
基因编辑技术的进步大幅提升了我们修饰人类基因组的能力。基于sgRNA介导的CRISPR- Cas9相关基因编辑技术能够在指定位点引入DNA断裂以失活基因功能或通过同源性DNA修复引导精确的DNA编辑,这些技术已针对基础DNA序列的靶向变化进行了优化,因此非常适合修复或引入致病性突变。然而,上述技术对内源性DNA修复机制的依赖提出了挑战,因为这些途径的复杂性可能使其难以进一步提升精确性。
最近的工作表明,表观基因组编辑有可能编写一种稳定的转录程序,该程序可以被人类细胞记住并传播,而无需可编程表观遗传调节剂的组成型表达。 此外,可以通过募集DNA甲基转移酶和KRAB结构域的混合物来沉默基因。然而,迄今为止,表观遗传记忆编写程序仅对少数内源性人类基因进行了沉默测试。此外,以前的可编程表观遗传沉默子设计为每个靶基因使用两个或三个融合蛋白,这在实验操作上很麻烦(尤其是对于多重基因靶向),从而使得基因靶向策略更加复杂。此外,基于TALE的KRAB与DNMT3A和DNMT3L结构域的融合导致基因沉默效率的长期低下。目前尚不清楚这些方法对于建立具有“遗传性”的基因沉默技术的通用性以及是否存在编写和维持可遗传表观遗传沉默程序所需的基因组特征。对此,一个合理的假设是:由单个失活Cas9融合蛋白组成的表观遗传编辑元件将有助于广泛地探索“可遗传”表观遗传基因沉默技术的实用性。
在最近发表在《Cell》杂志上的一项研究中,来自UCSF的Jonathan S. Weissman团队介绍了一种名为“CRISPRoff”的技术,包括其设计理念,开发过程和技术验证,CRISPRoff是一种可编程的表观遗传记忆编写器蛋白,可以持久抑制基因表达。研究发现,CRISPRoff的瞬时表达编写了一种表观遗传程序,从而维持人类细胞450多次的细胞分裂,突显出这种形式的基因沉默是稳定且可遗传的。 CRISPRoff表观遗传记忆可以使用称为CRISPRon的多部分表观遗传编辑器进行逆转,该编辑器可特异性删除DNA的甲基化修饰并募集转录元件。
使用全基因组CRISPRoff筛选,作者证明了这种方法可以持久且特异性地沉默绝大多数蛋白质编码基因,并且具有广泛的靶向性。此外,对于CRISPRoff介导的稳定基因沉默而言,规范的CpG岛标签不是必需的。
最后,作者证明了CRISPRoff可用于在人类干细胞中介导沉默增强和工程化的基因沉默程序,这些程序在神经元的不断分化过程中持续存在。总而言之,该系统使研究者们能够广泛探索表观遗传沉默的生物学规则,并提供了一个强大的工具来控制基因表达,靶向增强子以及探索表观遗传的原理。
3.Science:揭示蛋白QSER1保护DNA甲基化谷免受新生甲基化
doi:10.1126/science.abd0875; doi:10.1126/science.abh3187
DNA甲基化对哺乳动物的发育至关重要,它的失调可导致严重的病理状况,包括免疫缺陷-着丝粒不稳定-面部异常综合征(immunodeficiency-centromeric instability-facial anomalies syndrome, ICF)和小脑性侏儒症(microcephalic dwarfism)。酶DNMT和TET负责DNA甲基化的添加和去除,但它们如何协调调节甲基化景观仍然是一个核心问题。在一项新的研究中,通过使用一种基因敲入的DNA甲基化报告基因,美国研究人员在人胚胎干细胞(hESC)中进行了全基因组CRISPR-Cas9筛选,以发现DNA甲基化调节因子。相关研究结果发表在2021年4月9日的Science期刊上,论文标题为“QSER1 protects DNA methylation valleys from de novo methylation”。
这些作者重点研究了二价启动子(bivalent promoter),其定义为同时存在激活性(H3K4me3)和抑制性(H3K27me3)组蛋白标记,并且通常由多梳抑制复合物1和2(PRC1和PRC2)占据。在干细胞或祖细胞中,二价启动子被认为将发育调节因子维持在“蓄势待发的状态”,准备在分化时激活,并且它们对在功能失调的细胞背景(比如癌症或衰老)下的DNA高度甲基化敏感。构建一种基因敲入DNA甲基化报告细胞系提供了一个可视化观察表观遗传改变的绝佳机会,否则在干细胞状态下,基因表达变化是“看不见的”。通过使用PAX6 P0二价启动子作为代表位点,这些作者旨在在具有类似染色质特征的区域发现调节DNA甲基化的机制,这不仅可以了解发育过程中的基因调控,也可以了解疾病中的表观遗传失调。
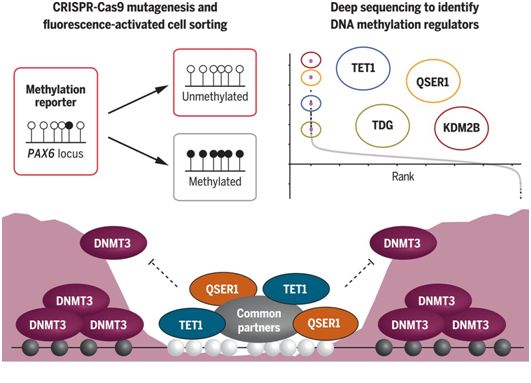
QSER1保护DNA甲基化谷免受新生甲基化。图片来自Science, 2021, doi:10.1126/science.abd0875。
DMV也被称为DNA甲基化峡谷(DNA methylation canyon),可识别存在于许多谱系的细胞中的较大(≥5 kb)的低甲基化区域,并在脊椎动物中保守。它们富含二价启动子、发育基因和转录因子,包括PAX6和HOX基因。QSER1和TET1在染色质免疫共沉淀测序(ChIP-seq)测得的基因组占有率中表现出高度的相关性,且两者在DMV中都高水平结合,然而新生甲基转移酶DNMT3A和DNMT3B在DMV中未结合,在DMV的两侧区域相对富集。进一步的蛋白质组和基因组分析显示,QSER1和TET1有许多共同的相互作用蛋白,相互依赖地高效招募到DNA上,并合作限制DNMT3A和DNMT3B在DMV中的侵袭。此外,剔除DNMT3B可以逆转QSER1基因敲除(KO)hESC细胞中的高度甲基化。此外,同时敲除QSER1和TET1对DNA甲基化和基因表达的影响比仅敲除QSER1或TET1都要强,并导致hESC不能分化为PDX1+NKX6.1+胰腺祖细胞。
4.Cell子刊:科学家通过结合CRISPR和人类ipsC技术,建立了人类白血病发生模型
doi:10.1016/j.stem.2021.01.011
来自美国纽约州西奈山伊坎医学院肿瘤科学系的Eirini P Papapetrou教授带领团队,在Cell子刊《Cell Stem Cell》杂志上发表了题为“Sequential CRISPR gene editing in human ipsCs charts the clonal evolution of myeloid leukemia and identifies early disease targets”的研究论文。
该研究团队专注于炎症信号传导,因为两种类型的早期变化分析主要由炎症相关基因决定。AML(SAR)阶段所需的基因表达和染色质可及性变化以及在早期(SA)阶段已经建立的基因表达和染色质可及性变化主要涉及炎症相关基因。此外,最早的持续基因表达和染色质可及性变化的分析主要涉及炎症相关基因。为了进一步支持这一点,可获得染色质的TF基序分析鉴定了NF-κB(先天免疫和炎症反应的主要介质)和FOS/JUN(AP-1),一种调节免疫和炎症反应的TF家族,作为主要的a和SA阶段的TF。最近的研究已经描述了来自TET2和DNMT3A突变的鼠模型的巨噬细胞中增加的炎症反应。该研究团队的数据表明,这些可能扩展到ASXL1突变,这是继TET2和DNMT3A之后的第三大最常见的CH突变,并且它们已经在HSPC水平上表现出来。该研究团队在A,SA和SAR细胞中HSPC水平的炎症信号传导失调的发现与MDS/AML HSPC中细胞内在先天免疫信号传导失调的最新证据一致。
多种独立的机制有助于MDS和AML中先天免疫信号通路的过度激活,它们聚集在涉及IRAK1,IRAK4和TRAF6的中心复合物上。包括该研究团队在内的临床前研究已经证明了用小分子或抗体策略抑制MDS和AML中IL-1R/TLR-IRAK-TRAF6信号传导的潜力。值得注意的是,IRAK1和IRAK4抑制剂目前正在进行AML,MDS,MPN,淋巴瘤以及炎症和免疫相关疾病的临床试验。此外,该研究团队的数据表明HSPC水平的细胞内在炎症信号传导失调已经存在于CH阶段。因此,该研究团队的数据支持开发针对这些途径的疗法的基本原理,这些疗法不仅针对MDS和AML,而且还可能针对CH阶段的早期干预。
总而言之,通过结合CRISPR和人类ipsC技术,该研究团队建立了人类白血病发生模型,使该研究团队能够绘制出构成疾病进展基础的分子事件图,并确定对AML至关重要且在疾病进展早期发生的分子变化,并且这可能为改善靶向治疗或疾病进展的生物标志物提供参考依据。
5.Mol Ther: CRISPR-Cas9基因编辑技术可用于治疗地中海型贫血症
doi:10.1016/j.omtm.2021.03.025
CRISPR/Cas9基因编辑技术被认为是治疗各种单基因遗传性疾病的最有希望的策略之一。在最近发表于《Molecular Therapy》杂志上的一项研究中,来自意大利费拉拉大学的Alessia Finotti教授等人首次通过CRISPR / Cas9基因编辑技术对β039地中海贫血突变进行了校正。结果证明,在对来自纯合的β039地中海贫血患者的红系前体细胞进行CRISPR / Cas9校正后,能够获得正常的β-globin。等位基因特异性PCR和测序证明了这一点。此外,校正后的β-珠蛋白mRNA的积累以及相关的β-globin和成人血红蛋白(HbA)也能够顺利生成。该结果为治疗地中海贫血症提供了新的思路。
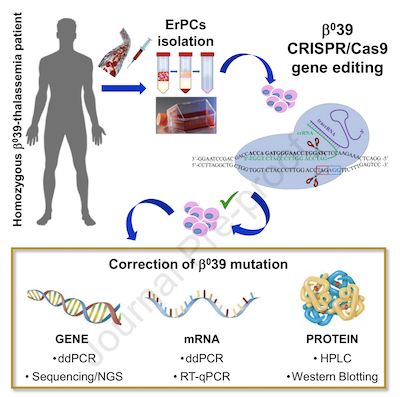
图片来自Molecular Therapy, 2021, doi:10.1016/j.omtm.2021.03.025。
基因组水平检测结果表明,CRISPR-Cas9能够高效地修正地中海性贫血患者中相关基因的突变。具体而言,β39突变核苷酸为胸腺嘧啶(T),经过编辑后成功地转变为正常的胞嘧啶(C)。丰度分析结果显示,在CRISPR-Cas9转染之后,至少4.5%的细胞中突变位点得到了修正。进一步,作者检测了修正后的mRNA表达水平。RT-PCR检测结果表明:修正后的mRNA表达水平相比未处理组中原始突变mRNA,至少高8倍左右。
最终,作者通过WB以及HPLC技术,验证了基因编辑后的细胞能够生成正确的β-globin,表明其功能能够得到恢复,此外,HbA蛋白的表达也得到了明显的恢复。
6.Mol Cell:光敏性sgRNA调控Cas9蛋白的失活
doi:10.1016/j.molcel.2021.02.007
对CRISPR-Cas9技术的精确调控可以提高其在基因编辑方面的安全性和适用性,然而这一目标目前却仍受 “不完全失活” 、 “速率过慢” 等缺点的限制。为了克服这些障碍,在最近一项研究中,来自约翰霍普金斯大学的Taekjip Ha教授等人设计了光敏感、可裂解的guide RNA(pcRNA),从而能够利用光照达到降解sgRNA分子,进而调控Cas9核酸酶基因编辑活性的目的。相关结果发表在最近的《Molecular Cell》杂志上。
首先,作者在gRNA分子中插入了光敏可切割基团,构建出了pcRNA分子,进而在体外实验中验证了该修饰后的sgRNA是否受光照的调节。实验结果表明,在缺乏光照的情况下,pcRNA与cas9混合物能够高效地切割DNA底物。相反地,在接受350nm光照的情况下,DNA分子则几乎没有受到切割。光照30s就足以达到灭活sgDNA-cas9复合体活性的目的。此外,作者还证明了pcRNA分子同时能够兼容其它类型的CRISPC-cas9平台,例如单核酸碱基编辑。在转染了AncBE4max以及pcRNA分子的HEK293T细胞中,光照2分钟就足以起到灭活单碱基编辑活性的效果。
众所周知,脱靶现象是基因组编辑技术中的难题,为了探究光敏基团修饰能否提高sgRNA-cas9靶向编辑的特异性,作者检测了基因组中VEGFA2以及HEK4等脱靶位点的胞嘧啶突变效应。结果显示,在共转染pcRNA与cas9/AncBE4max的HEK293T细胞中,脱靶位点的基因突变几率明显降低。与野生型sgRNA相比,光敏可切割的pcRNA的中靶/脱靶比例升高了2-9000倍。为了了解其背后的分子机制,作者通过体外切割实验检测了cas9/pcRNA的切割动力学特征。结果表明,在所有接受检测的靶向序列中,pcRNA的初始切割速率相比传统sgRNA明显更慢,但最终的切割速率达到相当的水平。此外,在sgRNA序列与靶向序列存在差异的情况下,传统的sgRNA仍旧能够介导高速率的切割,而pcRNA的切割速率明显更慢。这些结果表明pcRNA相比传统sgRNA具有更低的脱靶率。
之后,作者研究了pcRNA-Cas9达到有效切割活性时所需的最低时间,作者进行了系统的动力学分析并且绘制了时间曲线。结果表明,尽管因序列差异存在较高的异质性,但对于AncBEmax而言最低时间仅需4小时,而Cas9则需要36小时。
7.Nat Commun: "升级版"编辑器可在成年小鼠中进行基因校正并诱导癌症发生
doi:10.1038/s41467-021-22295-w
Prime Editor(PE)”是一类新型的基因编辑工具,该技术无需依赖于双链DNA断裂或外源供体DNA模板,即可介导基因组修饰。通过prime editing guide RNA中本身存在的“模板”序列,从而实现精确的单碱基替换或小规模的插入/缺失突变。为了探究该工具在成年小鼠中的基因编辑能力,来自麻省大学医学院的Wen Xue团队进行了深入研究。相关结果发表在最近的《Nature Communications》杂志上。
此前研究发现,Cas9元件中细胞核定位信号(NLS)序列的组成和数量会影响其基因编辑效率,原始的二代PE(PE2)包含两个NLS序列,转染以及细胞定位染色成像结果显示,约60%的蛋白质存在于U2OS细胞的细胞核中,约85%的蛋白质存在于HeLa细胞的细胞核中。在此基础上,作者在C末端添加N端c-Myc NLS序列,同时保留SV40 NLS序列,结果显示:改造后的PE2能够完全定位于细胞核内,作者将其命名为PE2*。此外,作者还对PE2*的主要框架进行了改造,使其能够识别更加广泛的PAM序列。为了研究改造后的PE2*能否提高其基因编辑效率。作者比较了HEK293T mCherry报告系统在分别转染了PE2和PE2 *之后的核苷酸转化比例。转染后3天,作者通过流式细胞术量化PE2以及PE2*的编辑效率。结果显示与PE2(9.2%至16.5%)相比,PE2 *的编辑效率提高了1.5-1.6倍(14.3%至26.4%)。在另外一个相似的报告系统中,PE2*的基因编辑效率比PE2提高了1.6-1.9倍。
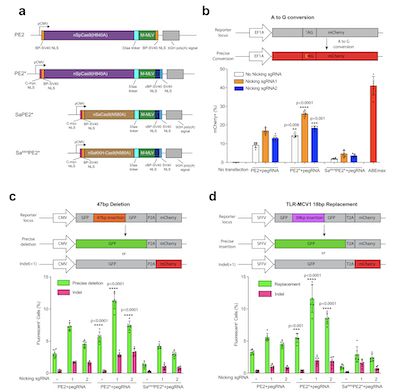
NLS序列改造提升PE2*基因编辑效率,图片来自Nature Communications, 2021, doi:10.1038/s41467-021-22295-w。
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
