
Cell Research:构建染色体融合小鼠模型、模拟染色体演化过程
来源:分子细胞卓越中心 2022-09-27 10:04
本研究证实着丝粒的断裂导致的染色体融合是染色体演化的原因、真核生物基因组组装的系统稳健性(Robustness)是染色体演化的重要基础。
Cell Research在线发表了中国科学院分子细胞科学卓越创新中心(生物化学与细胞生物学研究所)李劲松研究组撰写的题为Creation of artificial karyotypes in mice reveals robustness of genome organization的论文。该研究报道了基于类精子干细胞介导半克隆技术,通过CRISPR/Cas9靶向染色体重复序列,实现小鼠染色体融合改造,建立全新的稳定传递的染色体改造纯和小鼠品系,揭示了染色体融合的机制,并阐释了真核生物基因组组装的系统稳健性(Robustness)是染色体演化的重要基础。
染色体的稳定与变化是个体生存和物种演化的基础,也是遗传物质宏观调控规律的一体两面。染色体数目和结构的变异对个体造成不利影响,而新物种的形成往往又伴随复杂的染色体结构演化。系统生物学研究显示,距今约3到4百万年前,人与黑猩猩的共同祖先内部产生了染色体结构上的分异,两条独立的染色体通过头对头(类似罗氏易位,Robertsonian translocation)方式融合成为现代人的二号染色体(HSA2),这可能直接导致人类始祖与黑猩猩始祖之间的生殖隔离,成为人类物种进化的关键性事件。然而,这一事件发生的具体机制尚不清楚。
实验室常用小鼠(Mus musculus)的核型为40条染色体,在长期配繁过程中保持染色体数目和结构稳定,除Y染色体外,均为端着丝粒染色体(单臂染色体),这与人类染色体差异巨大。人类染色体中未发现端着丝粒染色体,而均以双臂染色体形式存在(包括近端着丝粒染色体)。然而,自然界中,野生小鼠存在较为广泛的染色体罗氏易位类型(端着丝粒染色体头对头融合)亚种,而罗氏易位类型分布不均衡,例如,Rb(2,4)(2号和4号染色体融合)融合类型和Rb(5,15)等融合类型在亚洲,欧洲美洲各地广泛分布,但Rb(1,13)和Rb(2,9)等融合类型尚未见报道。这引起了一系列思考,染色体融合如何发生?为什么小鼠的染色体演化存在端着丝粒偏好性?不同类型的染色体融合方式是否影响细胞和个体的生命活动?如何将小鼠的单臂染色体改造成更像人类染色体的双臂染色体?
2018年,中国科学家团队在国际上率先实现基于酵母的大规模染色体改造,利用头对尾的融合方式,将酵母染色体合并为一条,为染色体重排改造研究打开了一扇窗,但面对更复杂的哺乳动物,在个体水平改造染色体在技术上面临困难和挑战。在单基因突变和多基因突变遗传均可以高效建模的今天,攻克哺乳动物染色体结构变异建模是重要任务,而我国科学家独创的类精子干细胞技术为实现哺乳动物个体水平染色体改造投来了光亮。2022年8月26日,中科院动物研究所团队利用相似的头对尾的融合方式,获得了三只19对染色体的小鼠(Science)。
李劲松团队针对着丝粒核心基序Minor satellite(MinSat)设计了基于CRISPR/Cas9的靶向编辑方案,实现对着丝粒区域的靶向切割。将这样的着丝粒靶向切割系统转染到小鼠类精子干细胞中,可实现对小鼠两条染色体在着丝粒区域以“头对头”融合的方式发生罗氏易位,形成双臂染色体,模拟了自然界中在漫长演化过程中发生的染色体重排事件。研究在1128个转染着丝粒切割组件的单克隆细胞系中,建立了10株具有稳定的19条染色体的单倍体细胞系,其中有9株细胞系保持有基因组倍性平衡。
染色体着丝粒断裂融合形成了一个新的着丝粒。研究发现,所有和小鼠二号染色体(Chr2)发生罗氏移位之后形成的双臂染色体具有两个独立的着丝粒(MinSat富集区域)。进一步分析显示,小鼠Chr2与其他染色体不同,即在染色体末端具有两个着丝粒。双着丝粒染色体会造成在有丝分裂中存在不稳定性,易造成染色体断裂和基因组不稳定性。进一步分析发现,小鼠Chr2具有一个活化的着丝粒和一个失活的着丝粒。这暗示小鼠Chr2染色体和其他染色体相比,在进化历程中存在特殊的着丝粒形成事件。
染色体融合事件对于细胞的生命活动有何影响?研究对具有不同罗氏易位的细胞系进行转录组测序分析发现,携带单条罗氏易位染色体的“类精子干细胞”只具有最多不超过20个差异基因,这说明染色体“头对头”的融合方式对细胞表达谱扰动小,差异基因同时分布在参与融合和其余未参与融合的染色体上。
考虑到3D基因组结构对转录调控和进化都是重要的,研究考察了染色体融合对3D基因组结构的影响。罗氏易位细胞系和Wild-type细胞系相比,A/B compartment和TAD高度相似。此外,接近90%的染色质之间的显著相互作用可以在罗氏易位细胞系和Wild-type细胞系中同时被检测到。染色体融合直接拉进了两条染色体的物理距离,表现为两条染色体之间相互作用更加频繁。在着丝粒区域以及端粒区域,这种相互作用增强的趋势更加明显。研究还发现,小鼠Chr11与其余染色体相互作用在发生染色体融合后表现出较强的变化,即使其未参与到染色体融合事件。染色体融合对基因组结构和基因表达调控影响较小,这说明细胞内基因组结构及表达调控存在稳态调控,体现系统稳健性(Robustness)。
类精子干细胞具有替代精子使卵母细胞“受精”的能力,将染色体融合改造的类精子干细胞注入卵子后可以产生健康“半克隆”小鼠,并可以通过繁育获得携带纯和融合染色体的小鼠品系,在实验室条件下实现了哺乳动物的核型演化事件。该研究建立了四个独立的纯和染色体融合小鼠品系,雌雄纯和小鼠可自由配繁稳定传递融合染色体。
进一步,研究尝试了在单重染色体融合的类精子干细胞上进行叠加切割,产生了二重甚至三重染色体融合的类精子干细胞。随着染色体融合事件的叠加,细胞差异基因数目逐渐增加,这些基因同样分布在参与融合和未参与融合的染色体上,这暗示了染色体融合事件的累积带来转录组整体水平的扰动。多重融合的类精子干细胞作为父源遗传物质供体依然可以支持半克隆小鼠的产生,一步实现多重染色体融合小鼠模型的建立。
本研究证实着丝粒的断裂导致的染色体融合是染色体演化的原因、真核生物基因组组装的系统稳健性(Robustness)是染色体演化的重要基础。该研究为哺乳动物进行染色体结构的改造、动物新核型亚种的创造以及染色体结构变异疾病的模拟提供了可行的技术路线,开启了以小鼠为代表的哺乳动物染色体遗传改造的新领域。
研究工作得到科技部、国家自然科学基金、中科院及上海市科学技术委员会等的资助,并获得分子细胞卓越中心动物实验技术平台和细胞分析技术平台的支持。
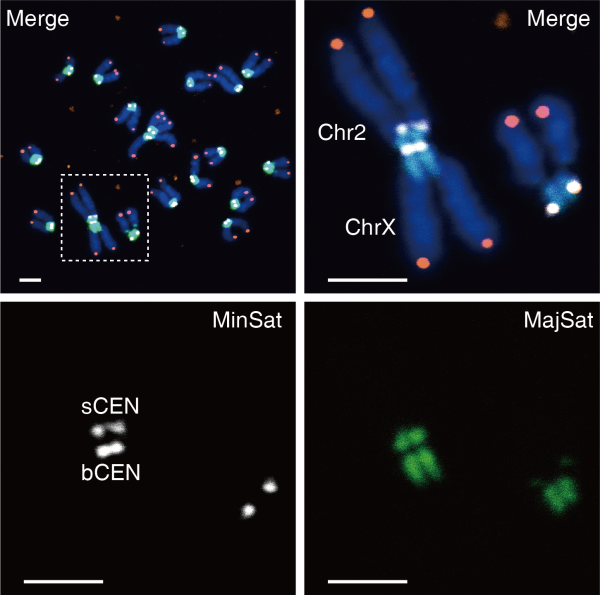
类精子干细胞中小鼠二号染色体和X染色体以头对头形式融合形成双臂染色体(红色:端粒;白色:着丝粒:绿色:近着丝粒)
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
