
号外:一看就懂的完整Wnt信号机制概览火热出炉!
来源:生物谷 2019-06-20 17:49
Wnt 配体蛋白是一类富含胱氨酸的糖蛋白,在发育、组织动态平衡、许多疾病的发生,包括癌症都起到了关键的作用。根据分子机制的不同,Wnt 信号传导通路分为三种:Wnt/β-catenin 信号通路、Wnt/PCP信号通路以及 Wnt/Ca2+信号通路。在哺乳动物中共有19种Wnt基因,其中部分基因包含选择性剪接异构体[22,35,38]。在功能上,不同的Wnt配体蛋白是高度保守的,由于不同Wnt配体
Wnt 配体蛋白是一类富含胱氨酸的糖蛋白,在发育、组织动态平衡、许多疾病的发生,包括癌症都起到了关键的作用。
根据分子机制的不同,Wnt 信号传导通路分为三种:Wnt/β-catenin 信号通路、Wnt/PCP信号通路以及 Wnt/Ca2+信号通路。
在哺乳动物中共有19种Wnt基因,其中部分基因包含选择性剪接异构体[22,35,38]。在功能上,不同的Wnt配体蛋白是高度保守的,由于不同Wnt配体表达模式的不同,会造成不同的Wnt配体在各个器官或组织发挥各自独特的作用[38]。在内质网合成Wnt蛋白的过程中,Wnt蛋白会被棕榈酰转移酶Porcupine棕榈酸修饰 [41],而在Wnt蛋白成熟的过程中,跨膜蛋白Wntless/Evi(Wls)和棕榈酰化的Wnt蛋白结合,并帮助Wnt蛋白运输到质膜[5,18,37,55]。
关于Wnt蛋白如何到达Wnt靶细胞存在两种假说,见图1,第一种,成熟的Wnt蛋白结合着Wls蛋白整合到分泌小泡或外泌体,位于分泌小泡或外泌体外侧的Wnt蛋白和Wnt受体相互作用 [14,28,34,44]。第二种,Wnt蛋白的转移是通过细胞间的FZD受体和跨膜E3 ligases Rnf43/Znrf3来介导的[11]。
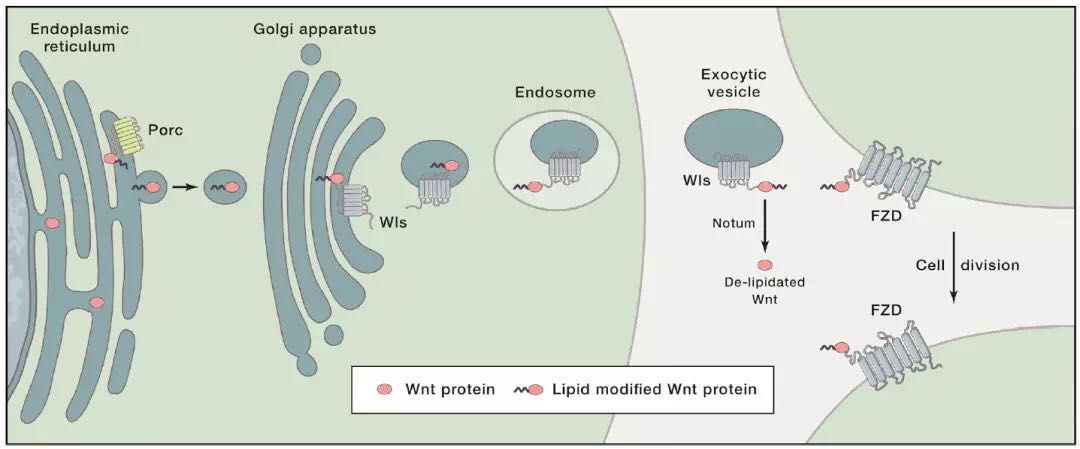
图1. Wnt蛋白分泌的模式 [38]
Wnt/β-catenin 信号通路
Wnt/β-catenin信号通路是依赖于β-catenin蛋白介导的一类Wnt信号通路,见图2。
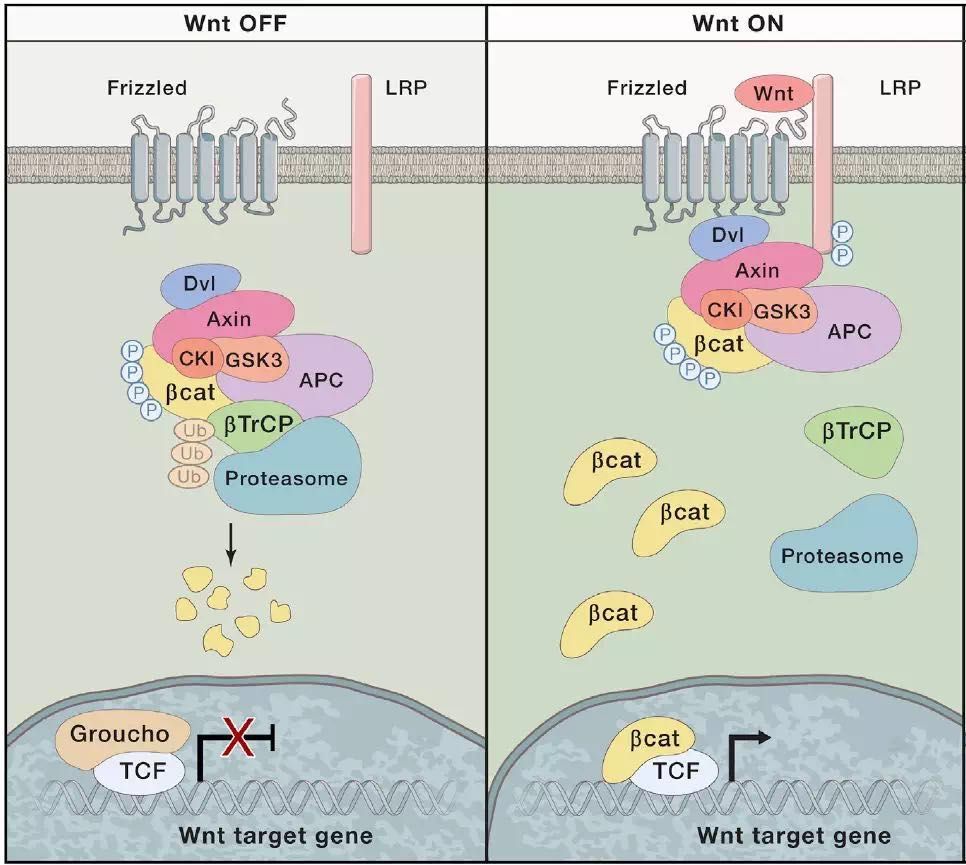
图2. Wnt/β-catenin信号模式图 [38]
Wnt/β-catenin信号通路由Wnt配体蛋白与七次跨膜蛋白FZD受体结合而启动,当Wnt配体蛋白会与靶细胞膜上的Wnt信号受体FZD和共受体LRP5或LRP6形成的异源二聚体相互作用[50],Dishevelled蛋白会被招募到细胞膜和FZD结合[7,25,29,52],而通过被招募到细胞膜上的Dishevelled蛋白和Axin相互作用,Axin-GSK3蛋白复合物也被招募到细胞膜处[50]。GSK3[51]和Cdk14(PFTK1)-Cyclin Y有丝分裂激酶磷酸化LRP5/6受体胞内段[8],随后,LRP5/6受体胞内段会进一步被CK1g磷酸化[9,56]。Dishevelled蛋白与FZD的结合提供了一个平台给予LRP5/6受体胞内段和Axin结合[38]。
胞质β-catenin是Wnt/β-catenin信号通路传导所必须的,而β-catenin的稳定性受到降解复合体的调控,APC、CK1a/d、GSK3a/b、Axin和β-catenin构成了降解复合体[38]。在没有Wnt配体到达靶细胞时,N端β-catenin先后被CK1、GSK3磷酸化[33],而磷酸化的β-catenin会被β-TrCP蛋白诱导泛素化随后被蛋白酶体降解[1,24],而当FZD和LRP5/6受体因Wnt配体刺激下形成Wnt-FZD-LRP5/6复合体时,β-catenin在胞质内的累积,这一过程目前存在着两种分子机制的假说:
第一种,降解复合体被重定位到细胞膜,而磷酸化的LRP受体可能直接抑制了GSK3的活性[49],在降解复合体中的磷酸化的β-catenin泛素化过程受阻,使得降解复合体被磷酸化的β-catenin所饱和占据,从而造成了新合成出的β-catenin在细胞质中的累积[2,32]。
第二种,Wnt-FZD-LRP复合体的形成动态调控了β-catenin的磷酸化[17],且使Axin蛋白去磷酸化,引起Axin和LRP以及β-catenin脱离,因而抑制了β-catenin的磷酸化[23]。
当β-catenin没有在胞质中累积时,TCF转录因子和Groucho蛋白结合转录抑制靶基因的表达 [6,42]。在胞质中大量累积了β-catenin后,β-catenin入核,Groucho/TLE转录抑制蛋白会被UBR5泛素化而失活[12],而β-catenin会和TCF结合并使其转变为一个转录激活蛋白,进而转录激活下游的靶基因,虽然其它的一些非TCF转录因子也被暗示参与介导了Wnt/β-catenin信号,但在哺乳动物细胞和果蝇中TCF作为最终的effectors介导了所有Wnt/β-catenin信号靶基因的直接激活[38]。
Wnt/PCP 信号通路
Wnt/ PCP 信号通路是一类不依赖于β-catenin的Wnt信号通路,见图3。
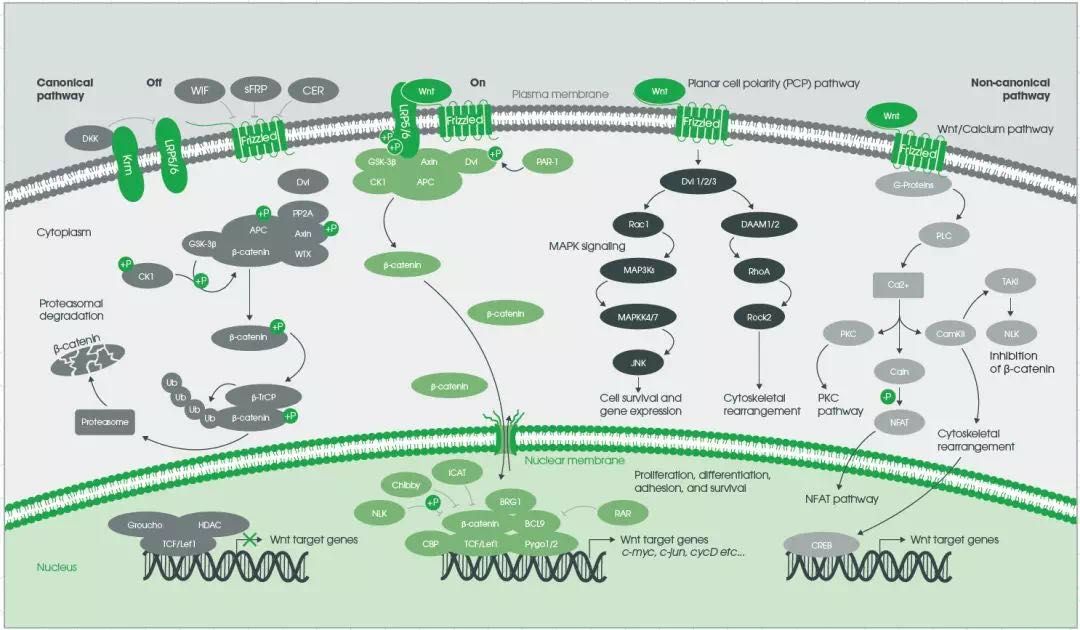
图3. Wnt信号总览模式图
PCP即平面细胞极性 Planar cell polarity,Wnt/PCP信号通路起始于Wnt配体和受体FZD结合,激活的FZD激活Dishevelled蛋白和G蛋白三聚体,并形成Dishevelled复合体的组装。而激活的Dishevelled激活小GTPases Rho和Rac。Rho 活化过程需要 Daam-1 参与,且Rho可同时激活 Rho-相关激酶 ROCK。Rac 活化则不依赖于 Daam-1,且可激活C-Jun N末端激酶 (JNK) ,因而调控了肌动蛋白细胞骨架和细胞粘附[15,20,46,47]。
Wnt/PCP 通路往往与细胞的极性,排列,迁移有关。毛发的平面极性,果蝇小眼或耳蜗中感觉毛细胞的排列,以及汇聚延展过程中的细胞迁移,都与Wnt/PCP通路有关[39,46]。
Wnt/Ca2+信号通路
Wnt/Ca2+信号通路也是一类不依赖于β-catenin的Wnt信号通路,见图3。Wnt蛋白和FZD受体结合后激活Disheveled,进一步通过G蛋白激活PLC,从而产生DAG和IP3,上调细胞内Ca2+。
另一方面,Disheveled的激活会活化cGMP特异的磷酸二酯酶PDE6,降低细胞内的cGMP,而使PKG失活,从而导致细胞内Ca2+浓度升高。Ca2+浓度升高会升高会激活下游通路,可能调控细胞粘附,或是抑制经典Wnt信号通路,或是激活CaMKII并通过NF-AT转录因子转录下游基因[10,30,36,47]。Wnt/Ca2+信号通路对细胞粘附及原肠形成中的细胞运动起着重要作用[15,26]。
Wnt/β-catenin 信号的负反馈调节机制
多种分子机制调节着Wnt/β-catenin信号的强度,见图4,包括在Wnt/β-catenin信号通路的配体,受体,胞内和细胞核层面[50]。

图4. Wnt/β-catenin信号负反馈调节模式图 [50]
在配体层面,Tiki蛋白切割N端的Wnt配体来抑制Wnt/β-catenin信号[57],而Notum蛋白去除Wnt配体蛋白的棕榈酸修饰也可以同样抑制Wnt/β-catenin信号[21,57]。而WIF或sFRPs蛋白可以通过直接和Wnt配体蛋白结合来阻止Wnt配体蛋白和受体复合物的结合[19,31,40,54],以此来达到抑制Wnt/β-catenin信号的效果。
在Wnt/β-catenin信号受体层面,Dkk1通过和Wnt配体竞争结合LRP5/6来抑制Wnt/β-catenin信号[3,48]。而Rnf43和Znrf3是一类跨膜E3泛素化连接酶,参与了FZD受体的内吞和蛋白降解,从而降低了Wnt/β-catenin信号强度[16,27]。
在Wnt/β-catenin信号胞内层面,Axin蛋白的水平增加了降解复合物的形成,因而促进了β-catenin的降解,以形成对Wnt/β-catenin信号的负反馈[13]。NKD1也在胞内层面抑制Wnt/β-catenin信号,其可能的机制是结合并抑制Dishevelled蛋白[43],而也有可能是NKD1直接和β-catenin蛋白相互作用并阻止其入核[53]。
最后在Wnt/β-catenin信号细胞核层面,神经系统中ICAT蛋白直接和β-catenin结合而抑制其在核内的活性,从而在具有时空性地抑制Wnt/β-catenin信号后向化的活性[45]。
Wnt/β-catenin信号通路与β-catenin有着密切的联系。
当缺少Wnt蛋白时,胞质β-catenin会被降解。β-catenin会被GSK3β和CK1激酶诱导磷酸化,磷酸化的β-catenin,Axin,APC和Disheveled会形成降解复合物,最终β-catenin被蛋白酶降解。
当细胞受到Wnt蛋白的刺激下,稳定化的β-catenin会进入细胞核,与转录因子TCF结合,激活下游诸多Wnt/β-catenin信号的靶基因,如Cyclin D1的转录。
Abcam现有两种Wnt信号通路抗体组合套装供您选择,欢迎点击产品货号查看详情:
ab242226,由5个与Wnt/β-catenin密切相关的重组兔单抗组成。
ab228526,由1支100μg 抗兔的HRP二抗和6支10μl的c-Myc,c-Jun,Cyclin D1,LEF1,c-Met 和TCF7重组兔单克隆抗体组成。
以上两种套装为您提供一个经济实惠的办法来研究Wnt信号通路的变化,满足您的实验需求!(生物谷Bioon.com)
参考文献:
[1] Aberle, H., Bauer, A., Stappert, J., Kispert, A., and Kemler, R. (1997). beta-catenin is a target for the ubiquitin-proteasome pathway. Embo j 16, 3797-3804.
[2] Azzolin, L., Panciera, T., Soligo, S., Enzo, E., Bicciato, S., Dupont, S., Bresolin, S., Frasson, C., Basso, G., Guzzardo, V., et al. (2014). YAP/TAZ incorporation in the beta-catenin destruction complex orchestrates the Wnt response. Cell 158, 157-170.
[3] Bafico, A., Liu, G., Yaniv, A., Gazit, A., and Aaronson, S.A. (2001). Novel mechanism of Wnt signalling inhibition mediated by Dickkopf-1 interaction with LRP6/Arrow. Nature cell biology 3, 683-686.
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
87%用户都在用生物谷APP 随时阅读、评论、分享交流 请扫描二维码下载->
