
Cell | 揭秘纤毛内运输的奥秘:逆向纤毛内运输的分子结构新发现
来源:生物探索 2024-08-05 09:00
该研究通过结合Cryo-ET和XL-MS技术,首次详细解析了逆向IFT列车的三维结构。揭示了逆向IFT列车的组装机制和蛋白质重排过程,为理解纤毛内双向运输的调控机制提供了新的视角。
近年来,通过冷冻电子断层扫描(Cryo-ET)和原位交联质谱(in situ cross-linking mass spectrometry, XL-MS),研究人员对顺向IFT列车的结构有了深入了解。然而,逆向IFT列车的结构以及顺向列车向逆向列车的转换机制仍未得到充分揭示。逆向IFT列车在纤毛顶端的重组过程涉及IFT-A和IFT-B复合物的全局重排,这对于理解纤毛内双向运输的调控机制至关重要。
7月26日Cell的研究报道“Extensive structural rearrangement of intraflagellar transport trains underpins bidirectional cargo transport”通过使用冷冻电子断层扫描和原位交联质谱技术,首次详细解析了逆向IFT列车的三维结构。研究发现,逆向IFT列车围绕IFT-A复合物形成对称聚合物结构,而这种重组过程需要顺向列车在纤毛顶端的完全解体。通过比较顺向和逆向IFT列车的结构,研究揭示了两者在货物结合位点的构象变化,这些变化确保了在双向运输系统中实现单向货物运输的精确调控。
该研究为深入理解IFT系统的工作机制提供了新的视角,并为纤毛相关疾病的研究和治疗提供了潜在的理论基础。纤毛的正常功能依赖于IFT系统的精确调控,而IFT系统的结构和功能异常会导致多种纤毛病,如多囊肾病(polycystic kidney disease)、Bardet-Biedl综合症和Joubert综合症等。因此,揭示IFT列车的结构和转换机制,不仅有助于基础生物学研究,还具有重要的临床应用价值。

在该研究中,研究人员利用了Cryo-ET和XL-MS技术,以揭示逆向IFT列车的三维结构。研究对象为温度敏感突变株 Chlamydomonas reinhardtii 细胞,通过对这些细胞进行温度处理后,制备了冷冻电镜网格(Cryo-EM grid)。
冷冻电镜网格制备:将处理过的Dhc1b-3细胞加入到已经进行等离子体清洗的Quantifoil R3.5/1 Au200网格上,然后加入胶体金溶液进行孵育。之后,通过液氮冷冻保存网格以用于后续的断层扫描。
Tomogram重建:使用MotionCor2进行运动校正,并通过CTFFIND4进行CTF估算。随后,通过imod/etomo管线进行半自动化的金颗粒对准,最终在etomo中重建断层图。
粒子挑选与子断层平均:在重建后的断层图中进行粒子挑选,并通过子断层平均技术提高信噪比,获得高分辨率的纤毛逆向运输列车的三维结构。
交联质谱分析:使用DSSO作为交联剂,对蛋白质进行交联反应。交联后的样品经过蛋白质组学处理,通过液相色谱-质谱联用技术(LC-MS)进行分析。所得数据通过ProteoWizard MSconvert转换为mgf格式,并在xiSEARCH中进行数据库搜索,最终使用xiFDR进行错误率估计和结果过滤。
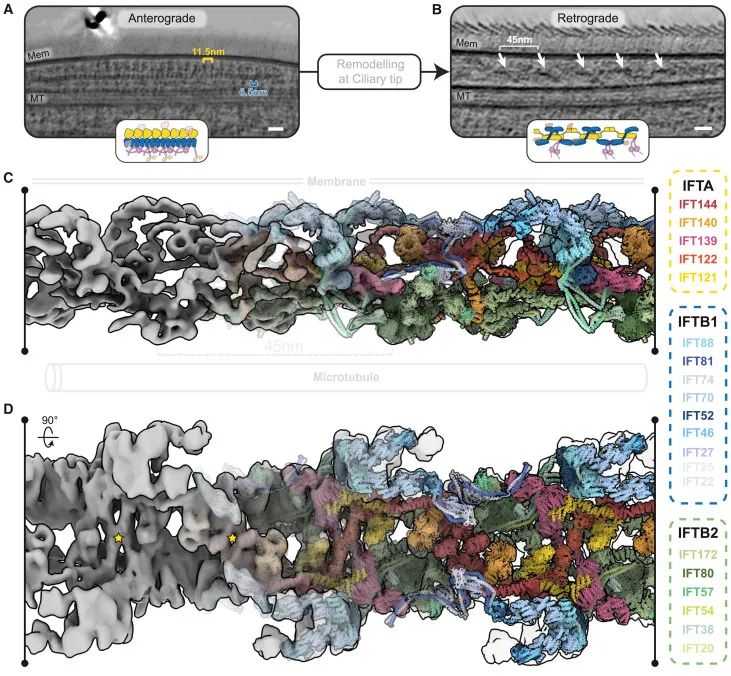
逆向IFT列车的分子结构及其关键特征(Credit: Cell)
逆向IFT列车的总体结构:逆向IFT列车围绕IFT-A复合物形成,对称排列。IFT-A复合物在逆向列车中起到了核心作用,形成了稳定的结构基础。IFT-B复合物在IFT-A复合物的周围排列,参与了列车的整体构建。
IFT-A复合物的具体作用:IFT-A复合物形成了中央的核心结构,通过IFT144与IFT121的相互作用形成了一个稳定的桥梁结构,这对于逆向列车的组装和功能至关重要。
结构特征和重复单位:逆向IFT列车显示出重复的结构单元,每个单元的长度约为45纳米。这些单元的排列方式确保了逆向列车的灵活性和适应性。
对称性:逆向IFT列车具有2倍对称性,这种对称性对于其在纤毛内的双向运输功能至关重要。对称结构使得列车能够在纤毛顶端实现从顺向到逆向的转换。
蛋白质相互作用:IFT-A和IFT-B复合物之间的相互作用是逆向列车形成的关键。IFT-A复合物的中心线沿着列车的长度延伸,而IFT-B复合物则在IFT-A的外围排列,形成了复杂的相互作用网络。
逆向列车的结构
研究发现,逆向IFT列车是一种高度灵活的聚合物结构,其核心为IFT-A复合物。
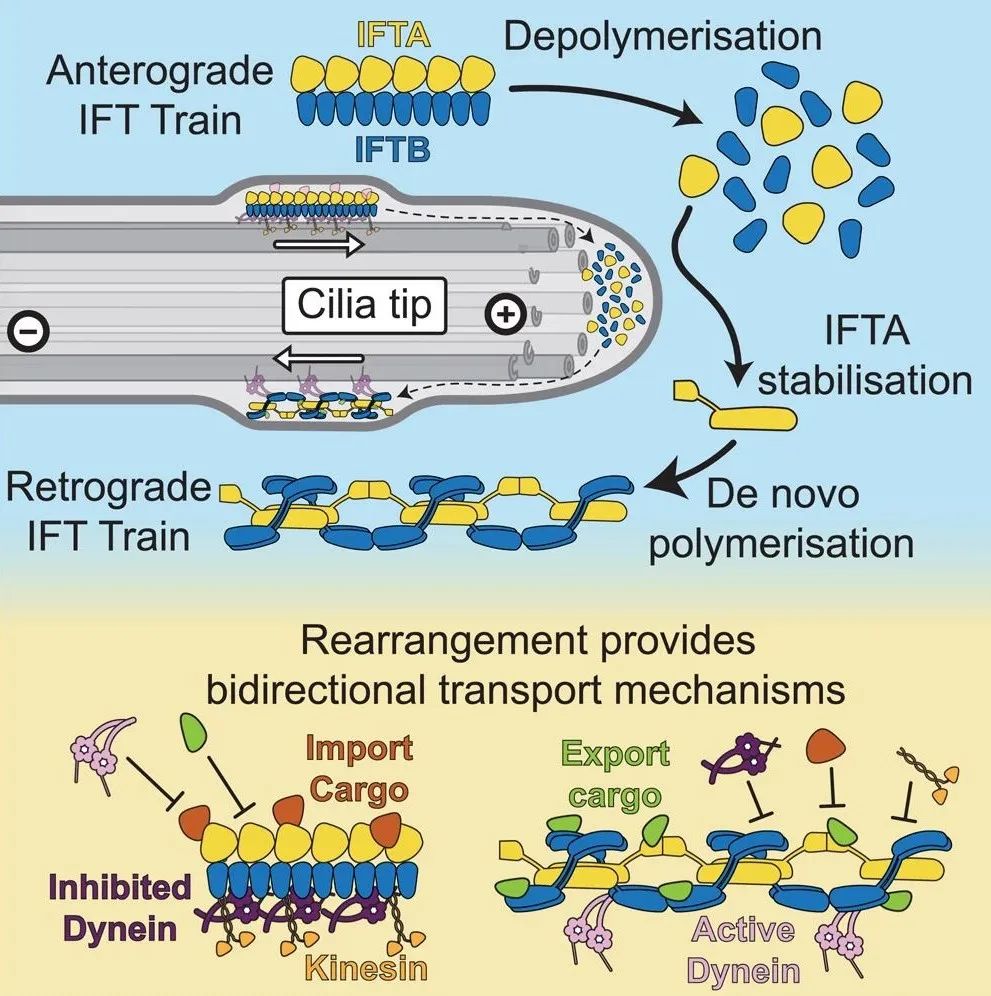
逆向列车的形成:在纤毛顶端,顺向列车进行完全解体,IFT-A和IFT-B复合物重新聚合形成逆向列车。通过电子显微镜图像分析,在600个野生型C. reinhardtii断层图中,研究人员识别出了95个逆向列车和741个顺向列车。逆向列车的平均重复距离为45纳米,相较于顺向列车的11.5/6.5纳米要长得多。逆向列车具有2倍对称性,其核心由IFT-A复合物组成,周围环绕着IFT-B复合物。
蛋白质重排:在逆向列车中,IFTB复合物的柔性结构域变得有序,以与IFT-A复合物相互作用。IFT144在其中通过形成桥梁结构,诱导逆向列车的形成。这种重排确保了逆向列车的稳定性和功能。通过对顺向和逆向列车的结构比较,研究人员发现顺向列车在纤毛顶端进行完全解体后,IFT144与IFT121形成了新的桥梁结构。这种重排使得IFT-A和IFT-B复合物在逆向列车中形成了新的相互作用界面,确保了逆向列车的稳定性和功能。
货物结合位点:逆向和顺向IFT列车在货物结合位点的构象上存在显著差异。例如,顺向列车中特有的货物结合位点在逆向列车中被遮蔽,确保了货物的单向运输。通过交联质谱数据分析,研究人员识别出了689个残基间的相互作用,其中包括48个IFT复合物蛋白的交联。这些交联在逆向模型中显示为合理的距离,进一步验证了其结构模型。这种结构变化确保了在双向运输系统中实现单向货物运输的精确调控。
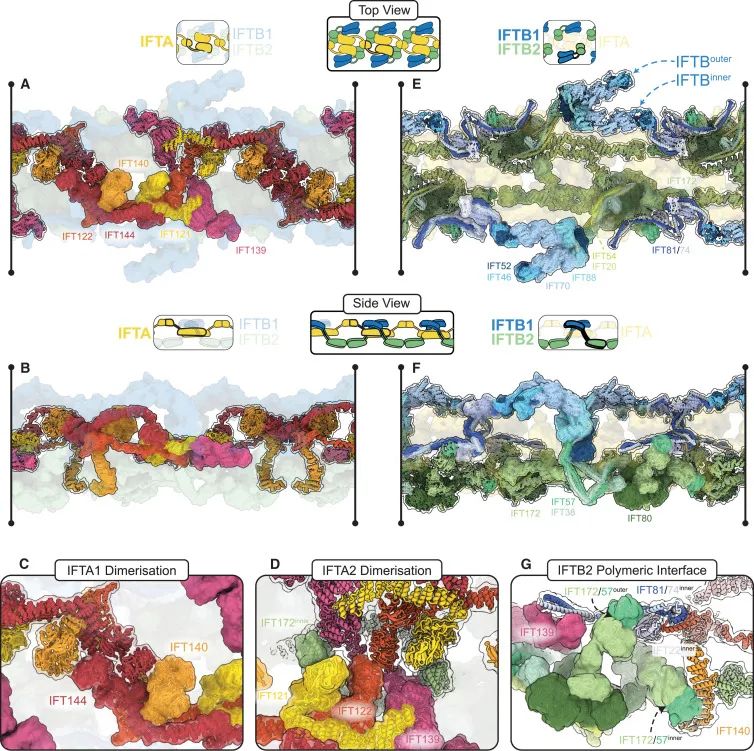
IFT-A复合物在逆向IFT列车中的核心作用和其形成C2对称结构的机制(Credit: Cell)
IFT-A复合物在逆向列车中的核心作用:IFT-A复合物(IFT-A complex)在逆向IFT列车中形成了一个连续的中心结构线。这个中心结构线是逆向列车的核心,贯穿整个列车的长度。
IFT-A复合物的对称性:逆向IFT列车展示了C2对称性(C2 symmetry),这意味着列车的两侧是对称的。IFT-A复合物在逆向列车中的排列方式使其具备这种对称性,确保了结构的稳定性和功能性。
IFT-A复合物的聚合界面:IFT-A复合物通过两个独立的同源二聚体界面(homodimeric interfaces)聚合。第一个界面由两个IFT144/140-WD亚基组成,它们相互缠绕形成“钩”状结构。第二个界面由IFT139和IFT122、IFT121-WD亚基组成,形成抗平行的二聚体结构。这些界面是逆向列车聚合的基础,确保了其稳定的双倍对称结构。
IFT-A和IFT-B复合物之间的相互作用:在逆向列车中,IFT-A复合物与IFT-B复合物之间的相互作用也得到了详细描述。IFT-B复合物包围在IFT-A复合物的外部,形成了列车的外“翼”。这种排列方式使得IFT-A复合物能够在中心形成核心结构,同时IFT-B复合物在外围提供稳定的支撑。
逆向列车的聚合机制:IFT-A和IFT-B复合物的排列方式不仅确保了逆向列车的稳定性,还通过特定的界面相互作用实现了列车的聚合。IFT-A复合物在逆向列车中的核心地位和其与IFT-B复合物的相互作用对于理解逆向列车的形成机制至关重要。
通过详细的结构分析,揭示了IFT列车在纤毛顶端的重组过程,并解释了其在单向货物运输中的作用。这些发现不仅深化了对纤毛功能的理解,也为未来的研究提供了坚实的基础。
该研究表明,IFT系统中的蛋白复合物在纤毛内经历了一系列严格调控的构象变化。这些变化不仅确保了纤毛的正常功能,也在细胞信号传递和感知中起到了关键作用。具体而言,顺向列车在纤毛基部组装,运输到纤毛顶端后完全解体,再重新组装成逆向列车。这一过程涉及复杂的蛋白质相互作用和构象变化。
尽管该研究通过Cryo-ET和XL-MS技术获得了逆向IFT列车的高分辨率结构,但这些方法仍存在一些局限性。例如,逆向列车的动态性和灵活性使得获取其全局结构信息具有一定挑战性。此外,未来的研究也可以进一步探索逆向列车在不同生理条件下的变化。
模式图(Credit: Cell)
综上,该研究通过结合Cryo-ET和XL-MS技术,首次详细解析了逆向IFT列车的三维结构。揭示了逆向IFT列车的组装机制和蛋白质重排过程,为理解纤毛内双向运输的调控机制提供了新的视角。这些发现不仅有助于基础生物学研究,也为纤毛相关疾病的研究和治疗提供了潜在的理论基础。
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
