
Nat Cell Biol:万刚团队揭示多相凝聚体结构的不混容性调节小RNA驱动的跨代表观遗传的机制
来源:生物探索 2024-10-05 14:47
该研究提供初步证据提示凝聚体中存在抑制不同凝聚体蛋白互作网络融合的调控蛋白,进一步的蛋白互作解析可以揭示多相凝聚体形成的具体分子机制。
生物大分子凝聚体(或称无膜细胞器)是细胞中主要由蛋白质或RNA通过相分离形成的通常具有液滴性质的基本组织单元,是有别于有膜细胞器的一类新型的细胞器,典型的凝聚体包括Stress granule、P body、核仁和生殖颗粒等等。这些凝聚体结构虽然没有磷脂双分子层,但是可以通过蛋白质和RNA形成表面与核心的分层结构 (如Stress granule)。此外,不同凝聚体还可以形成空间邻近但是不融合的多相结构,比如Stress granule和P body, 但是多相凝聚体形成的机制和生物学意义还很不清楚。
生殖颗粒是位于几乎所有后生动物生殖细胞核周的凝聚体,通过富集小RNA信号通路蛋白调控生殖细胞中RNA的命运和生殖细胞的正常发育。在秀丽隐杆线虫(C. elegans)中,生殖颗粒位于生殖细胞细胞核的核孔上面,并至少由四个空间上邻近且有序的凝聚体组成多相凝聚体 (P颗粒、Z颗粒、M小体、S小体)。不同的凝聚体富集不同但功能上相关的小RNA信号通路,如P颗粒富集线虫的piRNA相关蛋白、Z颗粒富集小RNA介导的跨代表观遗传相关蛋白、M小体富集小RNA扩增相关蛋白等。这些多相凝聚体被认为促进和协调复杂的RNA调控网络,异常的凝聚体结构通常导致RNA调控网络的紊乱、生殖细胞永生化的破坏和绝育。然而生殖颗粒的多相凝聚体形成的机制和协调小RNA信号通路并调控生殖细胞功能的机制还很不清楚。
2024年10月1日,中山大学生命科学学院万刚教授团队在Nature Cell Biology 杂志上发表题为HERD-1 mediates multiphase condensate immiscibility to regulate small RNA-driven transgenerational epigenetic inheritance的研究论文,发现了一个新的蛋白HERD-1,它可能通过维持生殖颗粒的多相凝聚体结构不混容性调节小RNA驱动的跨代表观遗传。

该研究首先通过优化的TurboID介导的生物素邻近标记技术,时空特异性地标记秀丽线虫生殖颗粒中不同凝聚体的蛋白并进行蛋白质谱分析,发现了大量未被报道的可能的不同凝聚体特异蛋白。生物信息分析表明不同凝聚体富集不同的蛋白质结构域和不同氨基酸基序的固有无序结构,提示这些结构域可以介导不同凝聚体的组装或者功能。通过功能筛选未被报道的编码PZM凝聚体组分的基因,发现了11个影响生殖细胞永生性和RNAi的基因,该功能筛选发现了一个同时参与这两个过程的新蛋白,HERD-1。令人意外的是,在HERD-1突变的线虫中,虽然生殖系RNAi介导的基因沉默缺陷,RNAi跨代表观遗传却增强到50代以上,同时观察到突变体中生殖颗粒的多相凝聚体结构被破坏而融合。结合亲和质谱和免疫共沉淀实验发现HERD-1可能通过降低或者阻止Z颗粒的蛋白网络与相邻两个生殖颗粒的支架蛋白(GLH-1和MUT-16)之间的相互作用,促进多相结构的形成。
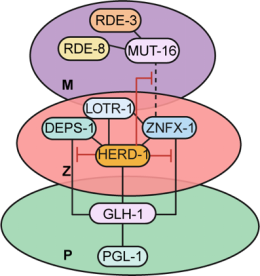
图一 HERD-1抑制多相凝聚体蛋白互作网络的模式图(Credit: Nature Cell Biology)
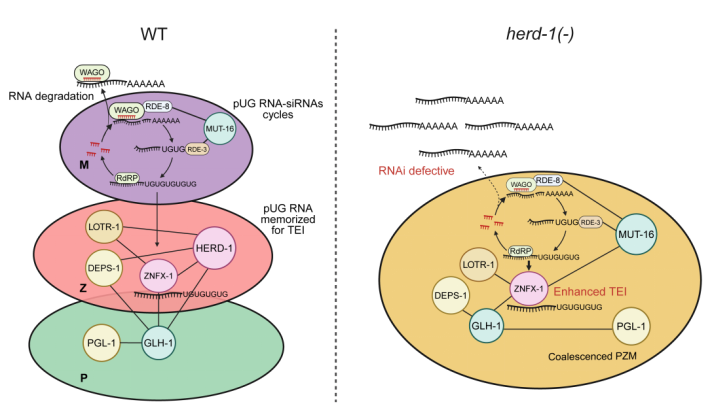
图二 HERD-1抑制生殖颗粒多相凝聚体融合和协调小RNA调控网络的模式图(Credit: Nature Cell Biology)
近年的研究表明,生物大分子凝聚体是蛋白质-蛋白质互作网络,非竞争的蛋白质的互作促进凝聚体的形成和凝聚体组分的招募,但是竞争性的蛋白质互作可以促进多相凝聚体的形成。
综上所述,该研究提供初步证据提示凝聚体中存在抑制不同凝聚体蛋白互作网络融合的调控蛋白,进一步的蛋白互作解析可以揭示多相凝聚体形成的具体分子机制。同时,这些结果提示多相凝聚体可以有序协调复杂RNA调控网络,而凝聚体的融合则可能会导致不同小RNA信号通路的竞争和RNA调控网络的异常,并最终导致生殖细胞永生性的破坏和绝育。
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
