
Cell:脑桥-髓质回路在快速眼动睡眠及帕金森病中的作用
来源:生物探索 2024-10-04 11:07
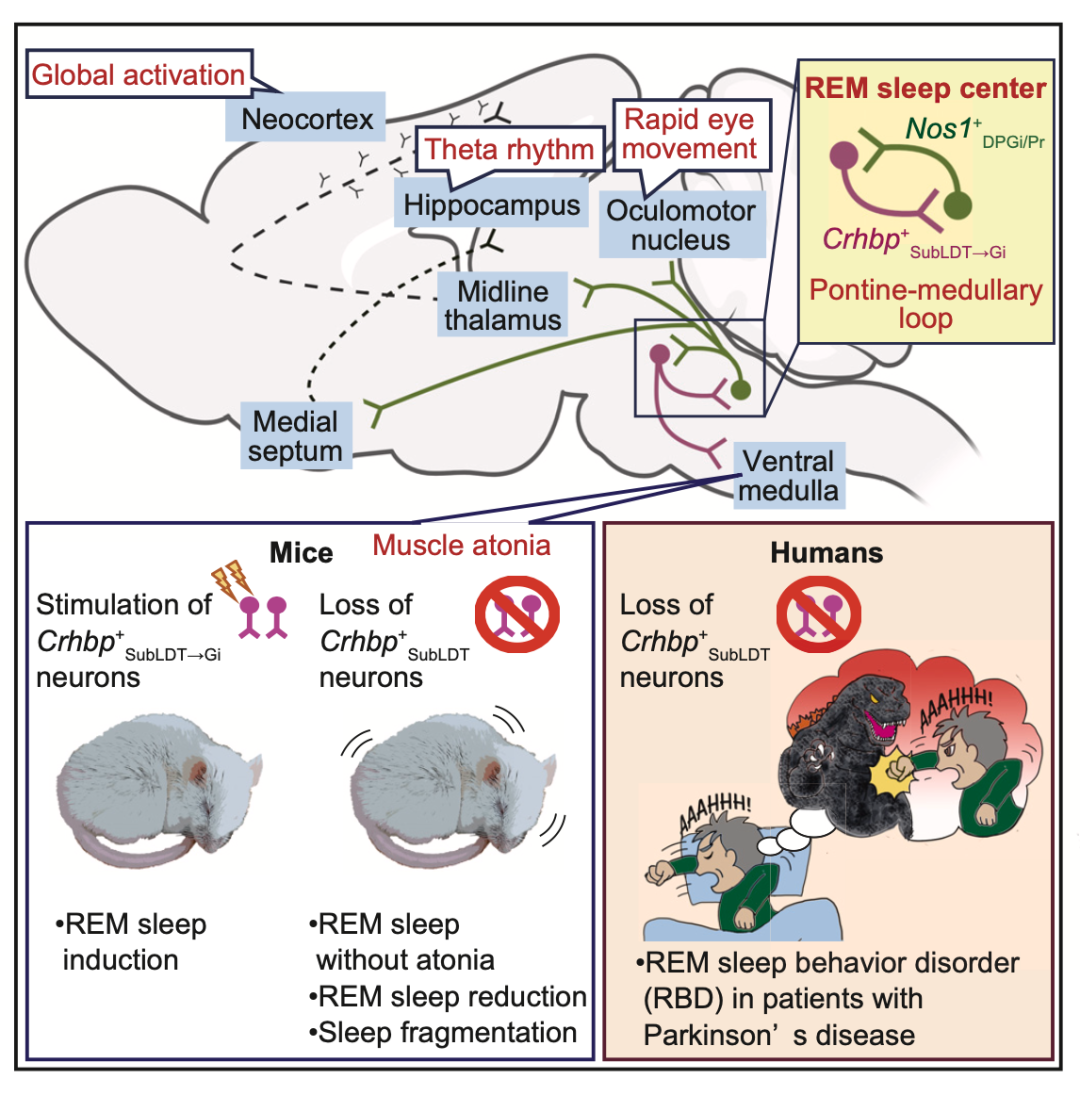
作者发现源自脑桥“Crhbp+神经元和髓质Nos1+神经元”神经回路可以诱导REMS,即使在清醒的小鼠中也是如此。
哺乳动物的睡眠包括快速眼动睡眠 (rapid eye movement sleep,简称REMS) 和非快速眼动睡眠 (NREM)。其中REMS也称作异相睡眠(paradoxical sleep)。NREM的特征是脑电图的高振幅慢波振荡,反映皮层神经元在去极化上升状态和超极化下降状态之间的同步转变。相比之下,REM的特点是皮质神经元的平均放电率增加,脑电图出现快速振荡和θ波以及骨骼肌活动的丧失(肌肉张力失调)等。脑干的脑桥被是调控REMS的关键结构【1】。
对猫的研究表明,蓝斑周围核 (peri-locus coeruleus,简称LC) -α是脑桥被盖(pontine tegmentum)内一个对诱发REM至关重要的区域【2-5】。随后的分析表明,啮齿动物的嗅下核 (sublaterodorsal nucleus,简称SLD) 与猫的LC-α是同源的【6,7】。在SLD内,抑制谷氨酸能神经传递可减少REMS量并缓解肌肉张力失调。该区域的谷氨酸能神经元是高度异质性的,而促进非快速眼动或觉醒的神经元则呈现混杂状态。虽然SLD中负责REM的神经元实体仍不清楚,但最近的研究揭示了其他区域的神经元在促进REM中的作用,包括下丘脑外侧的黑色素集中激素产生神经元,髓质腹侧的GABA能神经元和腹侧被盖区的多巴胺能神经元等。而这些发现表明,REM是由一个分散的、遍布全脑的神经元网络控制的,而不是一处特殊的中心环路。尤其值得注意的是,这些神经元的消融/慢性失活对REMS量的影响有限。相反,脑桥区的病变则会导致REMS几乎完全丧失【3,5】,这也表明SLD确实起着核心作用。因此,确定SLD中引起REMS的神经元亚型并揭示其神经环路是至关重要的。
近年来,REMS的缺陷,特别是REMS行为障碍 (REMS behavior disorder,简称RBD),因其与突触核蛋白病具有一定相关性,而引起了人们的广泛关注。RBD是一种睡眠异常症,其特征是在REM期间由于肌肉张力失调而导致梦境中的身体运动。队列研究表明,RBD是包括帕金森病 (PD) 在内的突触核蛋白病的前驱标志。除了RBD, PD患者还经常出现过度嗜睡和失眠,而PD患者睡眠不足的特点是REM和NREM的进行性下降。考虑到突触核蛋白病的晚期常伴有认知障碍,而记忆巩固需要REM,因此,阐明突触核蛋白病睡眠不足的原因是至关重要的。虽然在PD患者的脑桥被盖中观察到神经元变性和路易小体 (α-突触核蛋白的聚集体),但因为缺乏控制睡眠的神经元分子标记物,制约了它们在睡眠相关症状中的确切功能研究。
2024年9月19日,来自日本University of Tsukuba的Yu Hayashi研究组在Cell上发表题为A pontine-medullary loop crucial for REM sleep and its deficit in Parkinson’s disease的文章,发现一套源自脑桥和髓质的神经回路可以诱导REMS。

由于脑干神经元的异质性,识别快速眼动 (REM) 睡眠回路的特性及其与疾病的关系一直具有挑战性。在这项研究中,基于对脑桥被盖的转录组学分析,作者确定了一种特殊的神经元亚型,该亚型调节REMS/NREMS和REMS期间的肌肉张力。作者在小鼠身上发现,脑桥嗅觉下被盖中的表达Crhbp(corticotropin-releasing hormone-binding protein),并投射到髓质的神经元(也就是Crhbp+神经元)可以促进REM。而在接受Crhbp+神经元投射的髓质区域内,表达一氧化氮合酶1 的神经元(Nos1+神经元)投射到脑桥嗅觉下被盖,并促进REM,这表明脑桥和髓质之间存在一个正向的相互作用回路,并且,这一神经环路可以覆盖眼动睡眠回路。
Nos1+神经元也投射到控制广泛前脑活动的区域。切除Crhbp+神经元可减少睡眠并损害REMS。在伴有REMS行为障碍的帕金森病患者中,Crhbp+神经元的免疫响应水平大量减少,并含有病理性α-突触核蛋白,这为该疾病睡眠缺陷的机制提供了新的见解。
综上所述,作者发现源自脑桥“Crhbp+神经元和髓质Nos1+神经元”神经回路可以诱导REMS,即使在清醒的小鼠中也是如此。此外,帕金森氏症患者Crhbp+神经元缺失。这项工作建立了促进REMS的神经元遗传通路,为进一步研究其分布、上游和下游回路以及与人类睡眠障碍的相关性提供了理论基础。
模式图(Credit: Cell)
参考文献
1. Saper, C.B., Fuller, P.M., Pedersen, N.P., Lu, J., and Scammell, T.E. (2010). Sleep state switching. Neuron 68, 1023–1042.
2. Vanni-Mercier, G., Sakai, K., Lin, J.S., and Jouvet, M. (1989). Mapping of cholinoceptive brainstem structures responsible for the generation of par- adoxical sleep in the cat. Arch. Ital. Biol. 127, 133–164.
3. Sastre, J.P., Sakai, K., and Jouvet, M. (1981). Are the gigantocellular tegmental field neurons responsible for paradoxical sleep? Brain Res. 229, 147–161.
4. George, R., Haslett, W.L., and Jenden, D.J. (1964). A cholinergic mecha- nism in the brainstem reticular formation: induction of paradoxical sleep. Int. J. Neuropharmacol. 3, 541–552.
5. Sastre, J.P., Sakai, K., and Jouvet, M. (1979). Persistance du sommeil par- adoxal chez le chat apre`s destruction de l’aire gigantocellulaire du tegmentum pontique par l’acide kaı ̈nique. C. R. Seances Acad. Sci. D 289, 959–964.
6. Boissard, R., Gervasoni, D., Schmidt, M.H., Barbagli, B., Fort, P., and Luppi, P.H. (2002). The rat ponto-medullary network responsible for para- doxical sleep onset and maintenance: A combined microinjection and functional neuroanatomical study. Eur. J. Neurosci. 16, 1959–1973.
7. Pollock, M.S., and Mistlberger, R.E. (2003). Rapid eye movement Sleep in- duction by microinjection of the GABA-A antagonist bicuculline into the dorsal subcoeruleus area of the rat. Brain Res. 962, 68–77.
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
