
Cell:挑战传统感染模型——从HIV的“解体假说”到“完整穿越”
来源:生物探索 2025-01-23 10:15
“完整穿越”模型重新定义了HIV的感染逻辑,为研究人员理解病毒与宿主的攻防战提供了全新的视角。这一转变让人们更加接近破解HIV感染的核心机制,也为开发新的治疗手段开辟了道路。
艾滋病(HIV)感染一直是全球健康领域面临的重大挑战,而HIV-1病毒是导致这一疾病的主要元凶。尽管抗逆转录病毒疗法(ART)的广泛应用显著提高了患者的生存率,彻底消除病毒感染仍然面临科学难题。HIV-1病毒能够感染非分裂细胞(如巨噬细胞),并通过独特的机制将其遗传物质整合至宿主细胞的基因组中,从而在体内建立持久感染。1月17日Cell的研究报道“Passage of the HIV capsid cracks the nuclear pore”,发现这一过程的关键在于病毒独特的锥形衣壳(capsid)如何穿越细胞核孔复合体(nuclear pore complex, NPC),这一机制的复杂性和突破性为深入理解病毒感染打开了新视野。
该研究采用超高分辨率显微成像、冷冻电镜断层扫描(cryo-ET)以及分子模拟等技术,对HIV-1衣壳如何通过核孔的动态过程进行了深入探讨。值得注意的是,研究发现病毒衣壳在穿越核孔时不仅保持了结构的完整性,还会导致核孔复合体的环状结构发生破裂。这一发现挑战了传统的“衣壳在核孔外解体”假设,为病毒穿越宿主防线提供了全新解释。此外,研究揭示了衣壳与核孔蛋白(nucleoporins, FG-Nups)之间的特异性相互作用,这一过程可能通过增加核孔的空间灵活性来促进病毒入核。
该研究不仅为HIV-1病毒的核内转运机制提供了关键线索,还可能为未来的抗病毒疗法开发提供新思路,例如通过设计靶向核孔或衣壳相互作用的药物来阻断病毒的复制。这一突破性的研究为病毒感染机制的研究树立了新的标杆,同时也让我们更加接近破解HIV的“密码”。
HIV病毒的“破门”行动:它如何侵入细胞核?
自20世纪80年代发现艾滋病(HIV/AIDS)以来,这一疾病已成为全球范围内公共卫生的重大威胁。HIV感染者的总人数已超过3800万,每年新增感染病例依然数以百万计。尽管抗逆转录病毒疗法(ART)的发展显著延长了患者的生存期,但HIV病毒能够在宿主体内建立终身感染,且其变异性强、逃逸能力卓越,使得彻底治愈依然遥不可及。
在HIV病毒感染过程中,细胞核是一个至关重要的目标。核膜包裹着宿主的遗传物质,是病毒基因组整合的唯一入口。然而,核膜并非一堵静止的屏障,它由数千个核孔复合体(NPC)构成的微观通道网络组成。这些核孔直径仅约45至65纳米,既能选择性地运输必要分子,又有效阻止了大多数外来成分的进入。这一机制是宿主防御系统的重要组成部分。但令人惊讶的是,HIV病毒能够借助其锥形衣壳(cone-shaped capsid)突破这一复杂屏障,将其基因组成功送入细胞核内。
HIV-1衣壳的锥形结构是其突破宿主防御的关键“武器”。研究发现,这一衣壳由200-250个六聚体蛋白和12个五聚体蛋白拼接而成,形状既能保护病毒基因组免受宿主防御因子的攻击,又能与核孔复合体的苯丙氨酸-甘氨酸重复区域(FG-Nups)进行精准结合。数据显示,高达89%的病毒衣壳信号集中在核孔附近,其中17%的核孔同时与病毒衣壳直接相互作用,表明核孔是病毒穿越的关键“战场”。
而且,HIV衣壳不仅能够通过核孔直径与其宽端相近(约60纳米)的通道,还在此过程中保持其结构完整性。研究人员利用冷冻电镜和超分辨显微成像技术发现,病毒衣壳的窄端首先插入核孔,其形状逐渐引发核孔的扩张。这种扩张甚至导致核孔复合体的内环结构破裂,从而为衣壳的穿越腾出空间。这样的发现为传统认为“衣壳必须在核孔外解体才能入核”的假设提出了挑战。
锥形衣壳的秘密:它是如何进入核孔的?
HIV-1病毒的锥形衣壳(cone-shaped capsid)是一个复杂而精巧的分子装置。它由200-250个病毒衣壳蛋白(CA)六聚体和12个五聚体拼接而成,呈现出独特的锥形结构。这种形状不仅能够紧密包裹病毒RNA基因组,为其提供物理保护,还能为病毒在细胞内的运输和核内转移提供重要支持。研究显示,衣壳的宽端约为60纳米,与核孔的内径接近,而窄端的设计使其更容易插入核孔复合体(NPC)的中心通道。这种锥形设计堪称病毒突破宿主防御系统的“进攻利器”。
锥形衣壳的功能远不止是一个“防护壳”。首先,它是病毒反转录(reverse transcription)的主要场所。在衣壳内,病毒的RNA基因组被逆转录为双链DNA,从而为后续的基因组整合做好准备。同时,衣壳的存在还可以屏蔽宿主细胞内的DNA感受器,避免病毒在细胞质中被识别和清除。其次,衣壳与宿主细胞的微管马达蛋白相互作用,使病毒能够精准地被输送至核孔位置。更重要的是,衣壳还能作为一种“分子钥匙”,通过与核孔蛋白的特异性结合打开宿主细胞的核内屏障。
分子层面的“对接”:衣壳与核孔蛋白的结合
HIV-1衣壳成功进入细胞核的关键在于它与核孔复合体中苯丙氨酸-甘氨酸重复区域(FG-Nups)的精准结合。FG-Nups是核孔复合体的重要组成部分,通常会形成一种密集的“网状结构”,阻挡大分子通过。然而,研究发现,HIV-1衣壳蛋白的六聚体能够与这些FG重复区域特异性结合。这种结合不仅让衣壳能够附着在核孔上,还为其穿越核孔提供了足够的牵引力。
数据显示,在研究中观察到的所有核孔中,约有17%的核孔与病毒衣壳直接结合,而衣壳信号的89%分布在核孔附近,这进一步证明了核孔是衣壳穿越的核心场所。更为关键的是,这种结合还会触发核孔复合体的轻微扩张,为衣壳的通过创造物理空间。通过冷冻电镜断层扫描(cryo-ET),研究人员观察到衣壳的窄端首先插入核孔,并逐步“拉动”其宽端通过。
此外,分子模拟显示,衣壳与FG-Nups的结合产生了一种“抓取效应”,有效降低了其穿越过程中受到的阻力。这种效应让病毒能够以完整形态进入细胞核,而无需在核孔外解体。
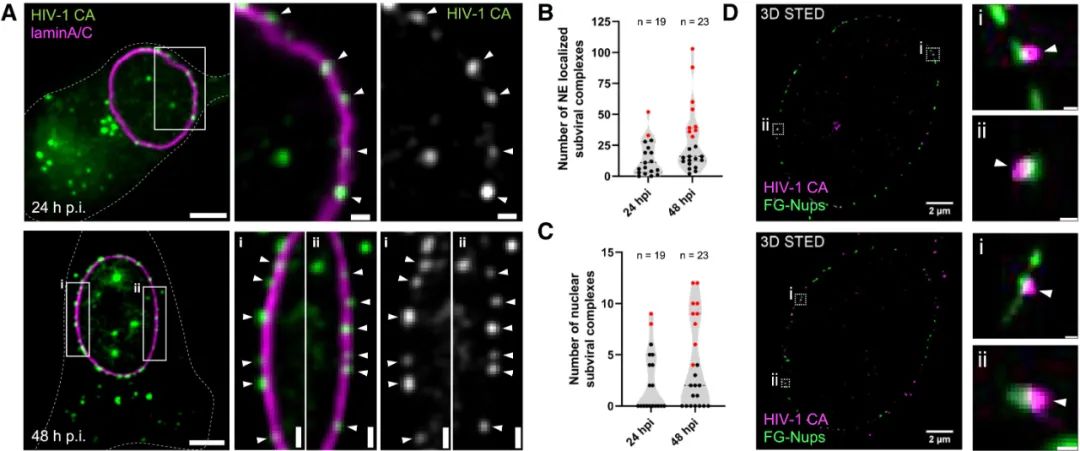
HIV-1衣壳在巨噬细胞核孔复合体(NPC)处的积累(Credit: Cell)
(A) 巨噬细胞中HIV-1 CA信号在核膜附近的分布
研究通过感染巨噬细胞样本(MDMs)并在感染后24小时和48小时进行标记观察,展示了HIV-1衣壳蛋白(CA,绿色信号)与核膜(由lamin A/C标记,洋红色信号)的相对定位。在共聚焦显微镜的z切片图中,可以清晰观察到HIV-1 CA信号在细胞核膜(NE)处显著积累。被lamin A/C信号定义的核膜区域显示出明显的CA信号聚集,尤其是在48小时后更为显著。局部放大的图像(标记为白色箭头)揭示了CA信号精确分布在核膜的定义边界处,这表明病毒衣壳与核孔复合体的关联。
(B 和 C) 定量分析CA信号的分布
(B) CA信号与核膜(lamin A/C)的共定位:图表显示,与lamin A/C染色共定位的CA信号数量在感染后不同时期的变化。结果表明,感染时间延长后,越来越多的CA信号与核膜区域共定位,这进一步验证了病毒衣壳在核孔复合体附近的积累趋势。
(C) 核内CA信号数量:分析了核内检测到的CA信号数量,结果显示感染48小时后,进入细胞核内部的CA信号显著增加。这表明病毒衣壳不仅在核孔处积累,还能够穿越核膜进入细胞核。
(D) 使用3D STED成像对病毒与核孔的精确关联进行深入解析
采用超分辨率的3D STED显微成像技术,研究进一步解析了HIV-1 CA信号(洋红色)与核孔蛋白(FG-Nups,绿色)的关联。在整个细胞核的3D重建图像中可以看到,CA信号直接位于由FG-Nups定义的核孔复合体区域附近或与之直接接触。局部放大显示,CA信号(白色箭头)清晰地与核孔复合体标记的结构直接相关联。这为病毒衣壳与核孔的物理交互提供了更直观的证据。
破裂的核孔:病毒穿越的代价
发现:核孔复合体的环状结构如何被“撕裂”
在HIV-1感染过程中,研究发现核孔复合体(NPC)并非被动地允许病毒穿越,而是经历了显著的物理变化。通过冷冻电镜和分子模拟,研究人员观察到病毒衣壳在穿越核孔时会导致NPC的环状结构出现“裂缝”。这种现象主要发生在NPC的内环区域(inner ring, IR),这是核孔的最狭窄部分,也是病毒通过时需要克服的关键物理障碍。
数据显示,约有一半的核孔在与病毒衣壳结合后出现了扩张,甚至裂解现象。这种“裂缝”表现为核孔的环状亚单位出现不对称排列,原本具有8重对称性的结构被破坏。通过分子模拟,研究还发现裂解的区域通常集中在衣壳接触和推进的位置,这表明裂缝的产生与衣壳的机械作用力密切相关。令人惊讶的是,尽管核孔结构受损,宿主细胞的整体功能并未因此受到严重影响。
动态过程解析:衣壳如何驱动核孔扩张
HIV-1衣壳是如何在分子层面实现对核孔的“挤压”的呢?研究显示,锥形衣壳的设计在这一过程中发挥了关键作用。病毒衣壳的窄端首先插入核孔中心通道,这一过程伴随着与核孔蛋白(FG-Nups)的紧密结合。随着衣壳的进一步推进,其宽端开始施加机械压力,促使核孔内环的亚单位向外移动。这种“扩张效应”让衣壳能够通过原本无法容纳其宽端(约60纳米直径)的核孔通道。
分子模拟进一步表明,这种扩张需要跨越两个主要的能量障碍:一是核孔蛋白的物理屏障,二是衣壳与FG-Nups之间的吸引力。在核孔扩张的过程中,这些能量障碍被逐步克服,而核孔的裂缝正是这一过程的标志性现象。
为什么破裂的核孔不会危及细胞生存?
核孔作为细胞的重要通道,其完整性至关重要。那么,为何核孔的裂解不会对细胞造成致命性损伤?研究人员提出了多个可能的解释。首先,尽管核孔的内环可能在病毒穿越过程中受损,但外围的核孔环和核膜结构依然保持稳定,这避免了细胞核的内容物泄漏。其次,裂解的核孔可能在病毒穿越后通过细胞内的修复机制迅速恢复功能,从而保障细胞的持续存活。
更有趣的是,研究发现HIV-1感染的巨噬细胞能够长期存活,并未表现出明显的核膜破裂或核孔完全丧失功能的现象。这表明,病毒可能在长期进化中找到了一种“恰到好处”的机制,既能成功突破核孔屏障,又能避免对宿主细胞造成过度损伤。对病毒而言,这种策略不仅提高了感染的成功率,还能为其在宿主内的长期潜伏创造条件。
核孔的破裂看似是一场“代价昂贵”的攻防战,但对于HIV-1来说,这是进入细胞核并完成感染的必要代价。而对于宿主细胞来说,这种微小而可逆的损伤,显然是一种可容忍的妥协。
如何“目击”病毒的穿越?
超高分辨率显微技术:探测衣壳行为
HIV-1病毒衣壳如何穿越核孔复合体(NPC)这一问题一直困扰科学界,而显微成像技术的发展为这一谜题提供了重要答案。研究人员利用超高分辨率显微技术,成功捕捉到病毒衣壳在穿越核孔时的行为细节。这种技术的分辨率可以达到10纳米以下,足以探测到衣壳的锥形结构以及它与核孔的相对位置。通过多通道荧光标记,研究人员实时观察了衣壳蛋白(CA)在核孔区域的信号强度变化,证明衣壳完整性在穿越过程中得以维持。
更重要的是,这种成像技术揭示了衣壳的动态行为,例如其窄端如何插入核孔并逐步推进至核内。这些发现为理解衣壳的“穿越路径”提供了直接证据,也证明了超高分辨率显微技术在揭示分子级别动态中的独特价值。
冷冻电镜与分子模拟:解密动态结构
尽管显微成像技术可以提供宏观视角,冷冻电镜(cryo-EM)与分子模拟则是解密病毒穿越过程中微观细节的利器。冷冻电镜能够在接近自然状态下捕捉病毒与核孔的相互作用,并生成高分辨率的三维结构图像。研究人员利用这项技术发现,HIV-1衣壳在进入核孔后,会引发核孔复合体的内环亚单位轻微移位。这种移位为衣壳的宽端通过创造了空间,同时核孔外围的环状结构保持稳定,防止核内物质的流失。
分子模拟则进一步揭示了衣壳与核孔蛋白(FG-Nups)之间的相互作用。模拟数据显示,衣壳蛋白表面特定的氨基酸序列可以与FG-Nups的苯丙氨酸-甘氨酸重复区形成短暂而高效的结合。这种结合不仅为衣壳提供了足够的“牵引力”,还触发了核孔的局部重构,为衣壳的通过铺平了道路。分子模拟还预测了不同亚单位的运动轨迹,揭示了核孔复合体扩张与回缩的协调过程。
这些技术手段的结合让研究人员获得了一系列关键数据。例如,研究显示,衣壳在核孔附近的停留时间大约为5-10秒,这表明衣壳通过核孔的过程是快速而高效的。此外,冷冻电镜断层扫描的图像显示,大约17%的核孔与病毒衣壳直接结合,而结合的衣壳中89%保持结构完整性。这些数据支持了病毒“完整穿越”的新模型,并否定了衣壳在核孔外解体的传统假设。
另一个重要发现是,裂解的核孔复合体在病毒穿越后能够迅速恢复,这可能与核孔复合体具有自修复能力有关。研究人员还通过对多个突变型衣壳的分析,验证了衣壳与FG-Nups的相互作用对病毒穿越过程的必要性。这些实验数据为设计干预策略提供了重要靶点,例如破坏衣壳与FG-Nups结合的小分子药物可能显著降低病毒的感染效率。
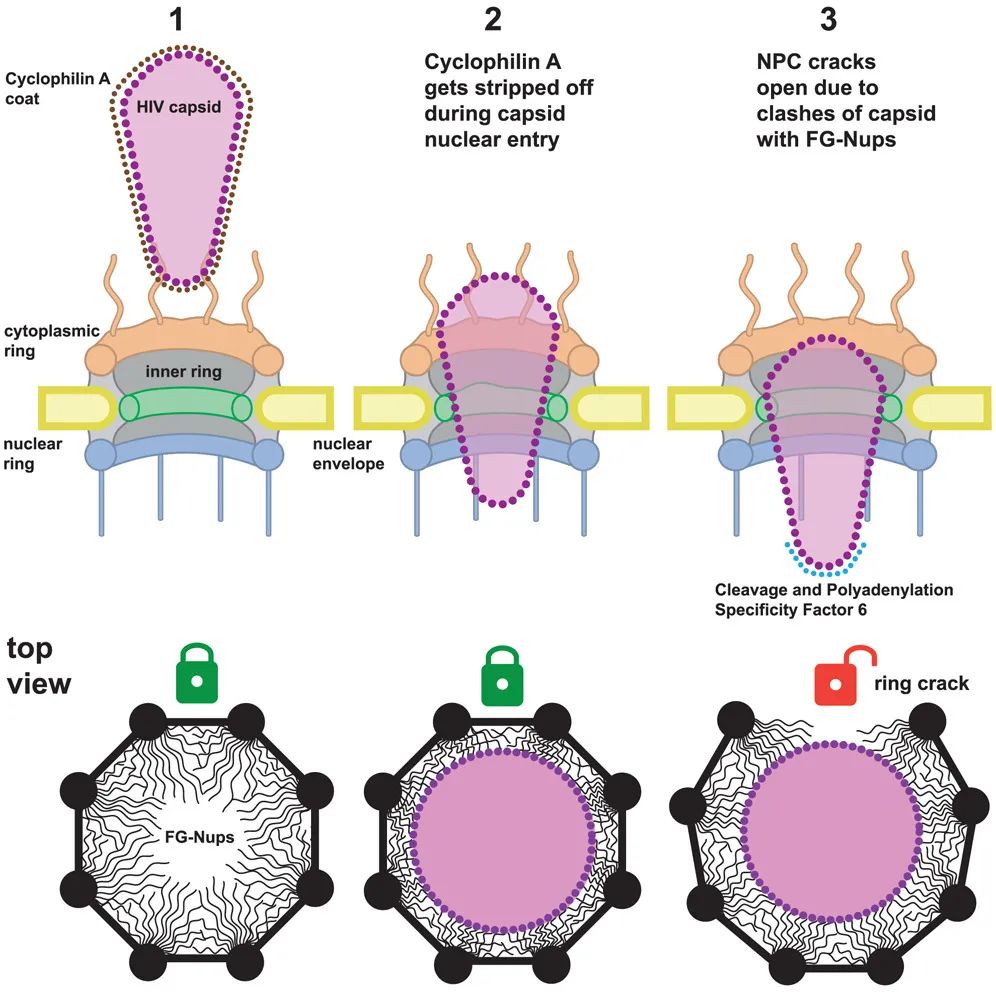
模式图(Credit: Cell)
挑战传统:从“解体假说”到“完整穿越”
在HIV-1病毒感染的研究中,传统观点普遍认为病毒衣壳在核孔外解体,其基因组裸露后通过核孔进入细胞核。这一“解体假说”基于早期对病毒和宿主相互作用的有限观察,认为衣壳的大体积无法通过狭窄的核孔。然而,这一假设存在明显的缺陷和争议。首先,裸露的病毒基因组在细胞质中易受到宿主防御因子的攻击,从而降低感染效率。其次,随着冷冻电镜和高分辨率显微技术的发展,越来越多的证据表明,病毒衣壳在核孔附近保持了相对完整的结构。
此外,“解体假说”无法解释病毒如何高效地将基因组成功导入细胞核,并避开宿主的免疫监控。这些问题促使研究人员对病毒的穿越机制进行重新审视,从而引发了对“完整穿越”模型的探索。
该研究提出了一种全新的“完整穿越”模型,即病毒衣壳无需在核孔外解体,而是以完整结构穿越核孔复合体。这一模型得到了多个实验结果的支持。例如,超高分辨率显微成像显示,衣壳在核孔区域保持完整,且其形状与核孔的尺寸具有高度适配性。此外,冷冻电镜断层扫描进一步证实,衣壳能够通过与核孔蛋白(FG-Nups)的特异性结合诱导核孔扩张,为其宽端的通过创造空间。
研究中一项关键发现是,病毒衣壳在穿越过程中表现出高度动态性。数据显示,核孔的内环亚单位在病毒穿越时会短暂移动,但整体结构在病毒通过后能够恢复。分子模拟分析还揭示,衣壳蛋白的特定氨基酸区域能够与FG-Nups形成暂时的高亲和力结合,这种结合产生的“牵引力”是衣壳完整穿越的动力来源。
新模型的提出不仅挑战了传统观点,还为病毒感染机制的研究提供了全新的理论框架。研究人员通过一系列实验发现,保持衣壳完整性对于病毒感染效率至关重要,而破坏衣壳与核孔的相互作用显著降低了感染能力。这些发现证明,“完整穿越”模型更符合病毒的实际行为。
“完整穿越”模型的提出从根本上改变了我们对HIV感染过程的理解。传统假设认为,病毒通过解体将基因组暴露给宿主,这一过程是被动的、低效的。而新的研究揭示,病毒具有高度精确的“入侵策略”,包括巧妙利用衣壳结构与宿主核孔的相互作用,从而实现基因组的高效传递。
这一发现还扩展了对病毒与宿主相互作用的认识。病毒并非简单地绕开宿主的防御,而是主动利用宿主的核孔复合体完成其生命周期的关键步骤。这一机制表明,核孔复合体的动态性和可塑性可能在病毒感染过程中扮演了比以往认为的更重要的角色。
对于科学界而言,“完整穿越”模型不仅推动了对HIV感染机制的深入研究,还为其他病毒的入核机制提供了参考。此外,这一研究为开发针对核孔或衣壳的创新疗法指明了方向。例如,设计能够干扰衣壳与FG-Nups结合的小分子药物,可能成为阻止病毒入核的新策略。
“完整穿越”模型重新定义了HIV的感染逻辑,为研究人员理解病毒与宿主的攻防战提供了全新的视角。这一转变让人们更加接近破解HIV感染的核心机制,也为开发新的治疗手段开辟了道路。
HIV研究的未来充满挑战,但也充满希望。从衣壳的结构特点到病毒与宿主的相互作用,研究人员正在逐步揭开这场微观攻防战的层层迷雾。理解HIV病毒的“秘密武器”不仅有助于攻克这一重大传染病,也为其他病毒感染的治疗研究提供了重要启示。
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
