
《细胞》:北京生命科学研究所曹鹏团队揭示“恶心-呕吐”反应的神经生物学机制
来源:奇点糕 2022-11-16 15:13
通过多种前沿的技术,曹鹏和他的团队发现位于肠道上皮的EC细胞可以感知肠毒素。EC细胞被激活后会分泌大量五羟色胺,表达五羟色胺3型受体的迷走感觉神经纤维捕获这一信号后被激活
大家都知道,摄入变质食物会引发恶心呕吐等一系列不愉快的反应。
恶心和呕吐都属于机体的防御反应,是为了避免病原体侵害身体。通过呕吐,机体可以促进有毒物质的排出;而恶心这一负面情绪,则使得我们可以避免再次摄入类似的有毒物质[1]。
恶心和呕吐,作为防御行为和情绪,其实都离不开神经系统的调节。那么,我们的大脑是如何感知肠道里的有毒物质,而后做出恶心和呕吐反应的呢?
之前的研究都试图通过有呕吐反应的动物模型(如犬类和雪貂)来解答这一问题[2]。这些研究发现恶心呕吐反应与肠-脑轴密切相关[3]。先前的研究还发现肠毒素和化疗药物顺铂可以激活脑干的孤束核,而迷走神经切除术可以有效阻断肠毒素和顺铂引发的恶心呕吐反应[4-5]。除此之外,药理学方法还证明了恶心呕吐反应与五羟色胺3型受体和速激肽1型受体有关[6]。
但是,由于在这些动物模型中很难实现分子遗传学操控,所以调控恶心呕吐反应的肠-脑轴的具体神经回路仍是不甚明了。
近日,来自北京生命科学研究所/清华大学生物医学交叉研究院曹鹏实验室通过小鼠模型解答了这个问题。这项工作得以发表在著名期刊Cell上[7]。
他们发现,毒素会激活肠道内的肠嗜铬细胞(EC),使得这些细胞释放五羟色胺,而五羟色胺会激活肠道的迷走神经。接着,迷走神经将信号传递给延髓背迷走神经复合体(DVC)中表达速激肽(Tac1+)的神经元,后者被激活后通过两条神经通路来分别调节小鼠的恶心和干呕行为。

论文封面截图
那下面就跟着奇点糕来看看曹鹏和他的团队是如何展开研究的吧。
基于小鼠模型在分子遗传学操控技术上的优势,他们首先成功地在小鼠身上建立了一个模拟食物中毒的模型,用来研究恶心和干呕反应的神经调控机制。
他们发现,金黄色葡萄球菌产生的肠毒素(SEA)可以导致小鼠表现出“大张嘴”的行为(retching-like behavior),也就是类似于干呕的行为;然后,通过条件性味觉回避实验(conditioned flavor avoidance, CFA),他们还证明SEA能引起小鼠表现出类似于“恶心”的厌恶性情绪。
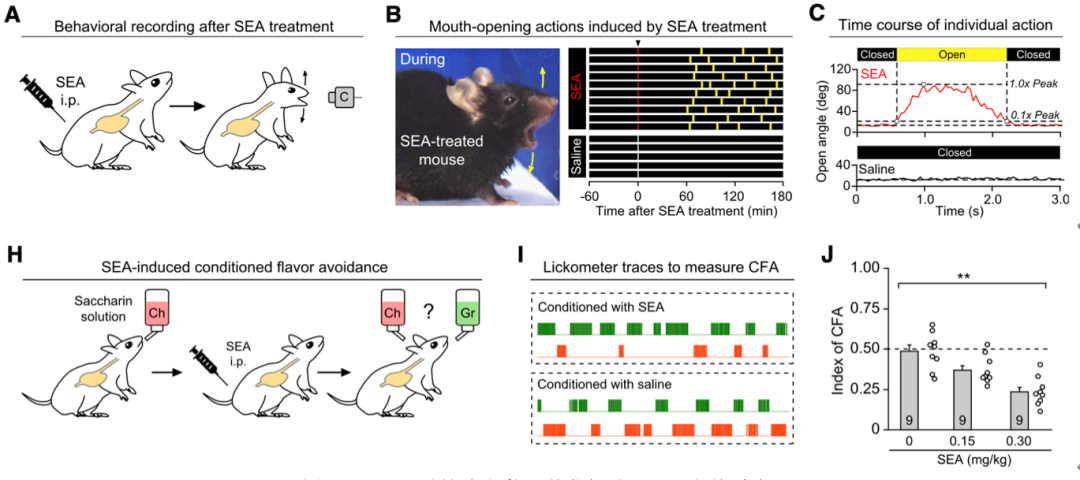
SEA可以导致小鼠呈现类似恶心和干呕的反应
由于之前的研究表明迷走神经与恶心呕吐反应密切相关,所以他们先对小鼠进行了迷走神经切除术,结果表明,迷走神经切除后的这些小鼠对于SEA的恶心干呕反应得到明显缓解。因为迷走神经投射的主要脑区之一是延髓中的DVC,包括孤束核(NTS)和极后区(AP),他们接下来探究了是否DVC对于SEA引发的恶心干呕反应有重要调控作用。
他们发现,DVC区域的神经元在SEA给药后被高度激活,而且特异性抑制这些神经元可以显著缓解SEA导致的恶心干呕反应。
那么,是DVC的什么神经元负责调控恶心干呕反应呢?
先前的研究表明DVC区富集有表达速激肽(Tac1+)的神经元,并且也有研究指明,如果抑制速激肽受体——神经激肽1型受体,可以有效阻断呕吐反应,因此作者们猜想是DVC区的Tac1+神经元负责调控肠毒素引发的恶心干呕反应。
为证明该猜想,他们通过化学遗传学技术特异性抑制了DVC区的Tac1+神经元,结果发现这可以显著抑制SEA引发的恶心干呕反应。
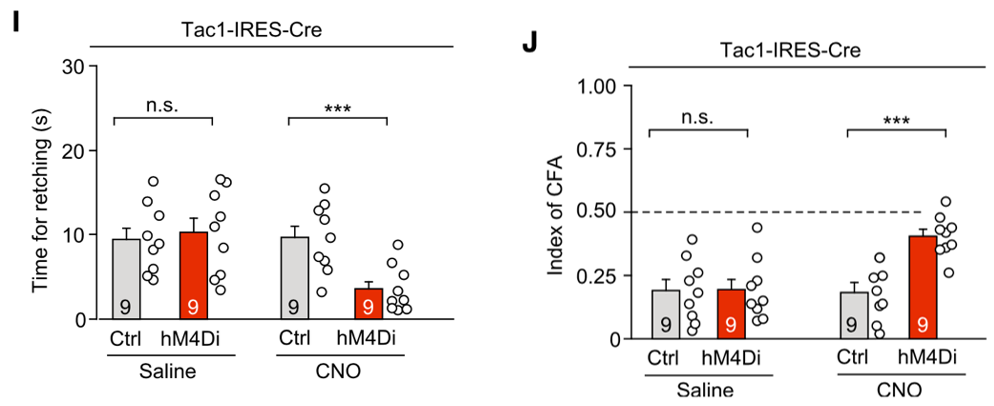
特异性抑制DVC区Tac1+神经元可以明显缓解SEA引发的恶心呕吐反应
另外还发现,这些Tac1+神经元分泌的Tac1相关神经递质和谷氨酸(glutamate)是这群神经元诱发恶心干呕反应所必须的。
下一个问题是,DVC的Tac1+神经元是如何获知肠毒素的信号的呢?
利用逆行示踪病毒标记DVC里的Tac1+神经元,作者们在胃窦和小肠的绒毛的黏膜末梢(mucosal endings)处发现了被标记的迷走神经纤维。在小肠里的黏膜末梢处,有一群细胞叫做肠嗜铬细胞(EC)。作者们惊奇地发现这些被标记的迷走神经纤维非常靠近于EC细胞。
因此,他们猜想这些EC细胞可以激活迷走神经元。
他们首先用染色的方法证明,大部分被标记的迷走神经元表达五羟色胺的受体—五羟色胺3型受体。接着他们发现,特异性敲除EC细胞中的五羟色胺可以明显抑制SEA引发的恶心干呕反应。也就是说EC细胞确实在SEA模型中激活了迷走神经元,而且这个通路是引发恶心干呕反应所必须的。
然后,作者们利用活体光纤钙成像技术测量DVC处Tac1+神经元在SEA模型中的反应活性,并且分别与膈下迷走神经切断术和HT3R拮抗剂处理组进行比较。结果显示,SEA模型中Tac1+神经元被激活依赖于迷走神经→DVC Tac1+神经元的信号传输。
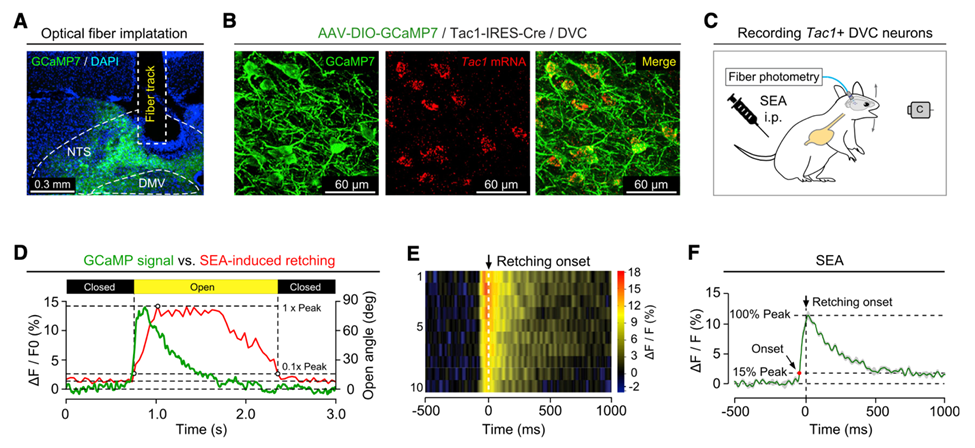
活体光纤钙成像技术证明SEA模型中DVC处Tac1+神经元被激活
那么,DVC的Tac1+神经元在SEA模型中被激活后,是如何引发恶心和干呕反应的呢?
为解答这个问题,作者们对DVC的Tac1+神经元进行了顺行示踪病毒追踪,他们发现这些Tac1+神经元主要投射到脑桥臂旁外侧核(LPB)、腹侧呼吸组吻端( rVRG)两个脑区。
他们还发现这两个神经回路几乎不存在重叠,也就是说在DVC有两个亚群的Tac1+神经元,它们分别投射到LPB和rVRG。有意思的是,作者们通过化学遗传学技术,发现这两个回路分别控制恶心和干呕两个反应:
①特异性激活DVC Tac1+→rVRG神经回路只引起小鼠的干呕样行为,但不诱发条件性味觉回避行为;
②特异性激活DVC Tac1+→LPB神经回路则不引起干呕样行为,却可以诱发条件性味觉回避行为。
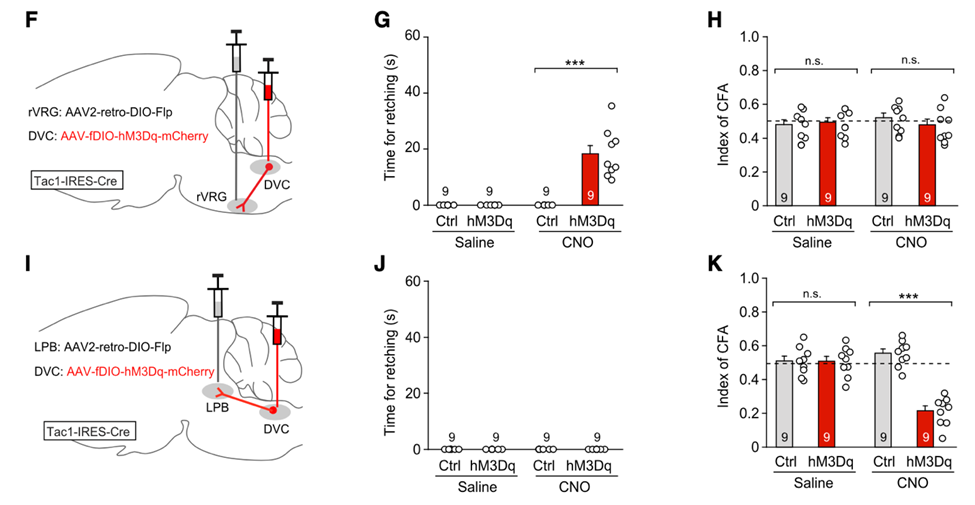
特异性激活DVC Tac1+→rVRG或DVC Tac1+→LPB神经回路对于SEA诱发的恶心和干呕反应的影响
除了食物毒素以外,化疗药物,比如顺铂和阿霉素,也会引发患者恶心干呕的行为。因此,作者们接下来探究上述神经回路是否也参与到了化疗药物引发的恶心干呕反应中。
他们发现,腹腔注射阿霉素能够诱发小鼠出现恶心呕吐的行为,而特异性抑制DVC Tac1+神经元、特异性敲除DVC区Tac1基因或者谷氨酸合成相关的基因,均能明显减弱这一现象。
另外,如果特异性抑制DVC Tac1+→rVRG回路,则可以特异性减弱阿霉素引起的干呕反应,而抑制DVC Tac1+→LPB回路则特异性减弱恶心行为。
综上所述,通过多种前沿的技术,曹鹏和他的团队发现位于肠道上皮的EC细胞可以感知肠毒素。EC细胞被激活后会分泌大量五羟色胺,表达五羟色胺3型受体的迷走感觉神经纤维捕获这一信号后被激活,进而将信号传入到位于延髓的DVC的Tac1+神经元。接着,Tac1+神经元将信号传输至LPB和rVRG两个脑区,分别引起恶心和干呕的反应。而上述的神经回路通路也参与到了化疗药物引发的恶心干呕反应中。
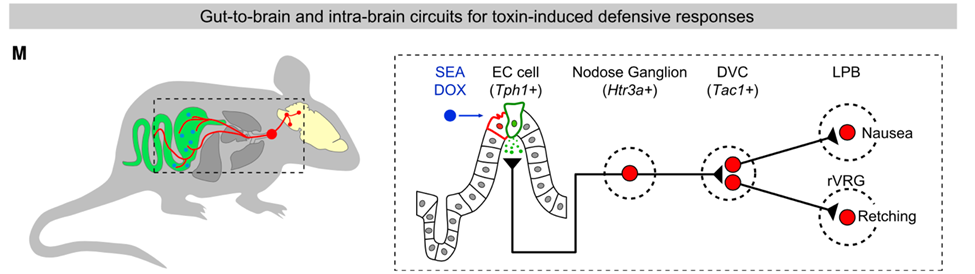
总结模型
此研究成果不仅加深了学界对肠-脑轴的认识,也为将来临床上对于恶心呕吐的治疗和干预提供了潜在靶点。
参考文献:
[1]. Horn CC. Why is the neurobiology of nausea and vomiting so important?. Appetite. 2008;50(2-3):430-434. doi:10.1016/j.appet.2007.09.015
[2]. Andrews PL, Horn CC. Signals for nausea and emesis: Implications for models of upper gastrointestinal diseases. Auton Neurosci. 2006;125(1-2):100-115. doi:10.1016/j.autneu.2006.01.008
[3]. Zhong W, Shahbaz O, Teskey G, et al. Mechanisms of Nausea and Vomiting: Current Knowledge and Recent Advances in Intracellular Emetic Signaling Systems. Int J Mol Sci. 2021;22(11):5797. Published 2021 May 28. doi:10.3390/ijms22115797
[4]. Reynolds DJ, Barber NA, Grahame-Smith DG, Leslie RA. Cisplatin-evoked induction of c-fos protein in the brainstem of the ferret: the effect of cervical vagotomy and the anti-emetic 5-HT3 receptor antagonist granisetron (BRL 43694). Brain Res. 1991;565(2):231-236. doi:10.1016/0006-8993(91)91654-j
[5]. Wang X, Wang BR, Zhang XJ, Duan XL, Guo X, Ju G. Fos expression in the rat brain after intraperitoneal injection of Staphylococcus enterotoxin B and the effect of vagotomy. Neurochem Res. 2004;29(9):1667-1674. doi:10.1023/b:nere.0000035801.81825.2a
[6]. Rojas C, Raje M, Tsukamoto T, Slusher BS. Molecular mechanisms of 5-HT(3) and NK(1) receptor antagonists in prevention of emesis. Eur J Pharmacol. 2014;722:26-37. doi:10.1016/j.ejphar.2013.08.049
[7]. Xie Z, Zhang X, Zhao M, et al. The gut-to-brain axis for toxin-induced defensive responses. Cell. 2022;185(23):4298-4316.e21. doi:10.1016/j.cell.2022.10.001
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
