
科研人员揭示多次跨膜蛋白的拓扑生成途径
来源:有机所 2024-06-02 11:39
该研究揭示了功能关键但与脂质相斥的pTMH拓扑生成途径,表明pTMH能引导膜蛋白翻译后拓扑结构重排,从而避免pTMH折叠时被暴露于疏水磷脂环境中。
在人类细胞内质网膜上合成的约5000种膜蛋白中,超过半数是多次跨膜蛋白。多次跨膜蛋白在细胞中作为离子通道、转运蛋白、受体蛋白、跨膜酶等,发挥着重要作用。这些功能多依赖于跨膜结构域的极性和带电氨基酸,而极性和带电氨基酸侧链通常具有排斥脂质的特性,导致其所在跨膜螺旋(TMH)具有较低的疏水性。统计表明,人类蛋白质组中约有30%的膜蛋白和超过50%的多次跨膜蛋白含有至少一个极低疏水性的TMH(pTMH)。在多次跨膜蛋白的成熟结构中,pTMH通常被周围的TMH保护,从而避免与磷脂双分子层的膜结构直接接触。然而,pTMH常常难以被转位子直接识别和插入。这些pTMH如何被识别并克服磷脂环境的疏水性、如何被包装进入成熟的多次跨膜结构,是相关领域尚未完全理解的科学问题。
近日,中国科学院上海有机化学研究所生物与化学交叉研究中心张在荣团队在《分子细胞》(Molecular Cell)上在线发表了题为AnATP13A1-assisted topogenesis pathway for folding multi-spanning membrane proteins的研究论文,揭示了由pTMH指导的多次跨膜蛋白拓扑结构生成途径。新生pTMH不能立即掺入内质网膜,而是穿越移位子中央孔进入水溶性内质网腔内,导致下游TMHs以与最终结构相反的错误朝向掺入内质网膜。合成结束后,P5-ATPaseATP13A1能识别并纠正“错误”的中间体构型,使得滞留于内质网腔的pTMH变得可识别,并整合、折叠进近成熟的结构,最终获得成熟构象。
以六次跨膜转运蛋白ABCG2和人类细胞作为实验模型,研究人员发现,ABCG2的新生pTMH2可以穿过转位子直接进入内质网腔,从而产生下游TMHs以错误朝向整合入膜的中间体构型状态。当位于羧基端的双赖氨酸翻译结束后,ABCG2的构型出现了几乎全局的拓扑重排过程。进一步研究发现,ATP13A1能感知这一双赖氨酸正电信号,当双赖氨酸被突变为负电或电中性氨基酸后,ATP13A1与ABCG2突变体的相互作用相较野生型大为减弱。敲除ATP13A1导致细胞内大量累积处于折叠中间体状态的ABCG2。ATP13A1能够在多次跨膜蛋白的拓扑结构成熟中发挥作用,促进了ABCG2中反向插入的TMH6从磷脂双分子层中解离。暴露在细胞质中的TMH6以正确的朝向重新插入内质网膜中,从而驱动上游TMHs的翻译后拓扑重排。
错误朝向的TMHs重排后,这种未成熟中间体能二聚化形成四级结构,可能促进后续pTMH2与其他跨膜螺旋束的组装,使pTMH2得以整合到膜中,形成pTMH2被其他TMHs所包围的最终结构。该研究揭示了功能关键但与脂质相斥的pTMH拓扑生成途径,表明pTMH能引导膜蛋白翻译后拓扑结构重排,从而避免pTMH折叠时被暴露于疏水磷脂环境中。
研究工作得到国家自然科学基金委员会、中国科学院和上海市科学技术委员会的支持。
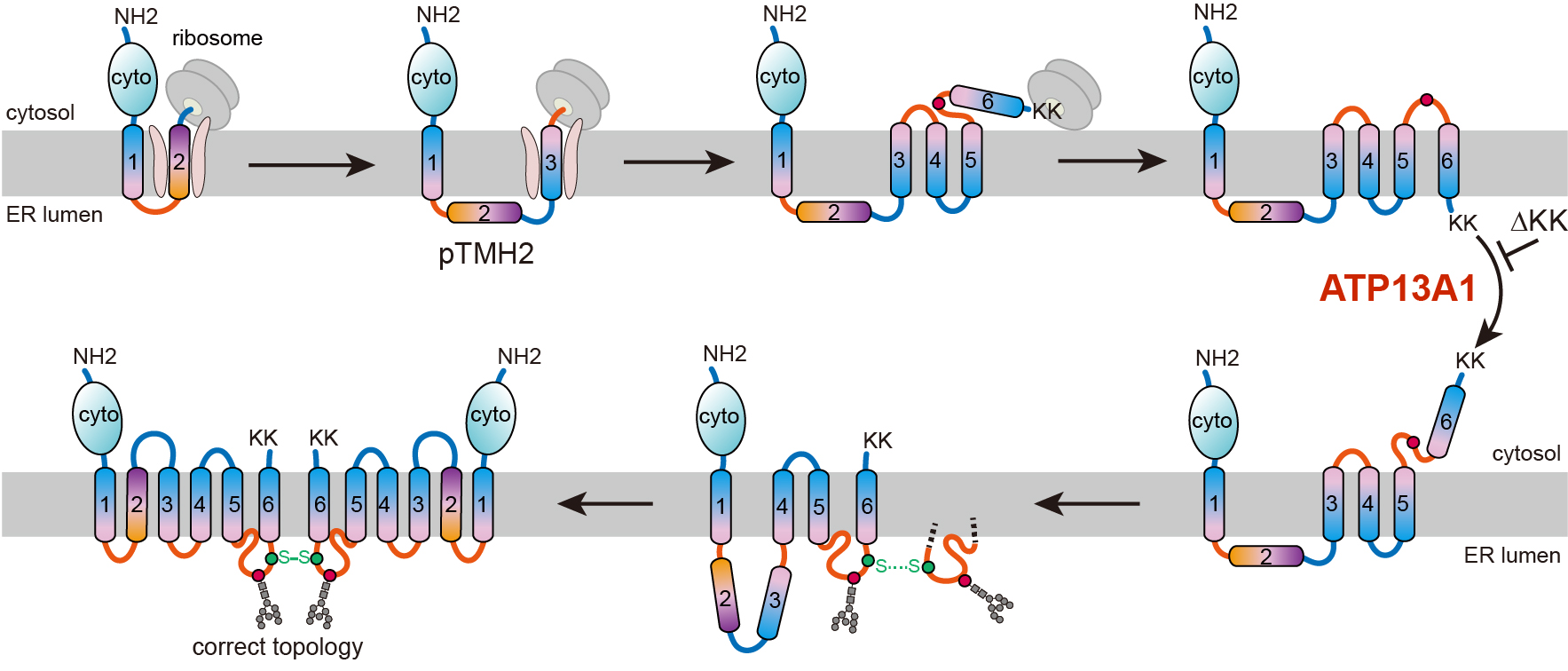
ATP13A1辅助ABCG2拓扑结构生成的途径示意图
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
