
《科学》子刊:科学家首次证实,新冠病毒S蛋白竟是溶酶体蛋白!
来源:奇点糕 2022-12-27 15:59
从细胞生物学的角度,Gould团队首次证实新冠病毒S蛋白是一种溶酶体蛋白,在β冠状病毒感染的细胞中,除了受体结合、膜融合以外,还能改变溶酶体的结构并重塑溶酶体的功能。
新冠疫情爆发至今已经三年,虽然奥密克戎致病性有所减弱,但新冠病毒仍在快速变异,而变异主要集中在S蛋白。因此,破解S蛋白核心变异对新冠病毒进出细胞的影响,对新冠病毒溯源、抗体疗法、疫苗设计尤其重要。
近日,由约翰·霍普金斯大学医学院Stephen Gould教授领衔的研究团队,在著名期刊Science Advances发表重要研究成果【1】。该论文的第一作者是郭晨栩,通讯作者是Stephen Gould,其他研究人员包括蔡尚睿、Yiwei Ai、Maggie Li、Eduardo Anaya、Andrew Pekosz和Andrea Cox。

论文首页截图
通过研究新冠病毒S蛋白在细胞内的转运,Gould团队发现S蛋白是一种溶酶体蛋白,2020年初出现并迅速占据主导地位、成为所有毒株共有的D614G突变并不是对人类宿主的适应。相反,D614G的出现纠正了由早期弗林蛋白酶切割位点插入(FCSI)突变所引起的一系列S蛋白功能缺陷,包括S蛋白向溶酶体的运输。
Gould团队2020年底曾在预印平台biorxiv上刊登过相关研究结果【2】,是世界首篇提供明确证据表明S蛋白能够重塑溶酶体结构和功能的论文,进一步支持以溶酶体为核心的β冠状病毒进出细胞的机理模型,并揭示了D614G突变作为基因内抑制因子,在新冠病毒进化、免疫、预防、治疗等方面的重要意义。

论文第一作者郭晨栩(左一)、通讯作者Stephen Gould(右一)和团队成员蔡尚睿(右三)、Yiwei Ai(左二)等
S蛋白是抗击新冠病毒最关键的蛋白质,因为它识别并介导受体结合,促进病毒进入细胞,依然是绝大多数疫苗的唯一抗原和抗体疗法的首要目标,而且它还一直在快速变异。
不同于其他SARS相关冠状病毒,新冠病毒在其起源之时或之前获得了FCSI突变,使S蛋白能被弗林蛋白酶介导的加工预先激活,以便其他丝氨酸蛋白酶如TMPRSS2进行切割,为新冠病毒提供了一种新的、不依赖于胞内体/溶酶体的入侵机制去感染细胞,从而使新冠病毒能够通过呼吸系统快速、有效地传播。
尽管FCSI增加了新冠病毒可以采用的感染机制、可以识别的受体范围、可以感染的细胞类型以及可以传播的途径和效率,但也引起了S蛋白结构的显著变化。由于改变结构的变异几乎总会伴随一些不利的副作用,FCSI会带来有害的特性也就不足为奇了。具体而言,FCSI导致内体/溶酶体感染途径出现重大缺陷,新冠病毒在缺乏TMPRSS2表达的VeroE6等细胞系中复制不良,在连续传代后,迅速出现FCSI逆转突变体【3-7】。
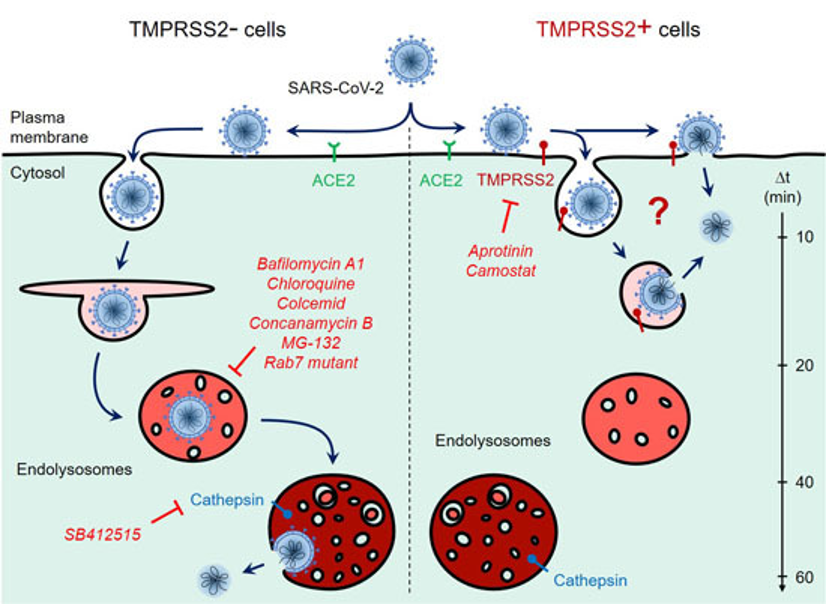
新冠病毒感染宿主细胞的两个途径【7】
疫情开始不久,新冠病毒便获得了另一个有利的变异(D614G突变)。这是新冠病毒在人群中传播后出现的第一个变异,也是后来所有毒株共同拥有的变异。它显著增强了新冠病毒的感染力、传播性和病毒载量,奇点网此前也曾多次报道。
一般来说,病毒会朝着减少其细胞表面表达的方向进化,这可能是因为适应性免疫反应定向选择了能够更好地逃避免疫监视的突变病毒。为了检测D614G是否能够减少S蛋白在细胞膜上的表达,研究人员把新冠病毒原始毒株(WT)和D614G突变株的S蛋白克隆出来,分别整合到一个由四环素操纵子调控的基因表达体系中,并通过两个不同的对照实验(生物素细胞表面标记结合亲和素亲和层析分离,以及细胞表面荧光标记流式细胞术)发现,D614G使细胞膜S蛋白的表达量降低至原来的三成水平,而这两种S蛋白的总表达量相似。
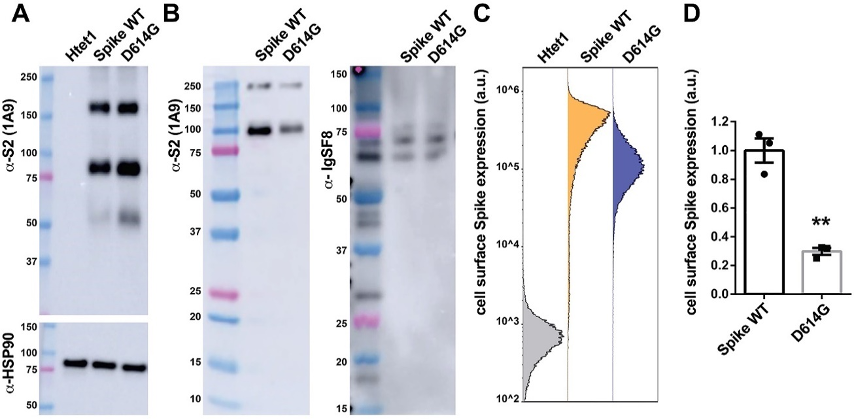
D614G导致细胞表面S蛋白表达量显著下降
众所周知,蛋白质在细胞内的转运是一场零和博弈。鉴于S/WT和S/D614G的总表达水平相似,既然D614G使其在细胞膜的表达量减少了70%,那么它一定是被运输到了细胞内部。
研究人员通过分析早期新冠患者的血浆,利用不同患者产生的抗体,追踪S蛋白在细胞内的转运。果然,他们在表达S/WT的细胞中发现了强烈的细胞表面信号,而在表达S/D614G的细胞中找到了更多的细胞内部定位。有趣的是,S/WT显示出大量与高尔基体标记物GM130荧光共定位,而S/D614G则高度集中在多个大型非高尔基体细胞器中。
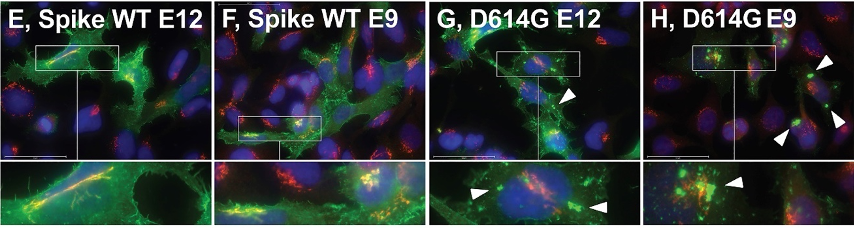
D614G导致大量S蛋白被运输到细胞内部的非高尔基体细胞器中
此前曾有多则报道,β冠状病毒能将宿主细胞的溶酶体转化为病毒储存和释放的容器,所以新合成的β冠状病毒颗粒不是通过经典分泌途径释放的,而是被运输到溶酶体并在那里不断积累,直到被溶酶体胞吐作用大量释放。
为了确定S/D614G高度集中的细胞器是否就是溶酶体,研究人员做了一系列实验,包括免疫荧光显微、免疫金电子显微技术等,证实了(1)S蛋白是一种溶酶体蛋白并能引起溶酶体聚集,(2)D614G将S蛋白从细胞膜重定向到溶酶体限制膜上,(3)与S/WT相比,D614G导致更多的S蛋白(约1.5倍)转运到溶酶体上。
研究人员接下来测试了D614G在新冠病毒感染的情况下,是否也会影响S蛋白在细胞内的转运。他们分别用原始毒株和D614G突变株感染Vero E6/TMPRSS2细胞,用免疫荧光显微镜和流式细胞术等方法分析,得到了相似的结果。
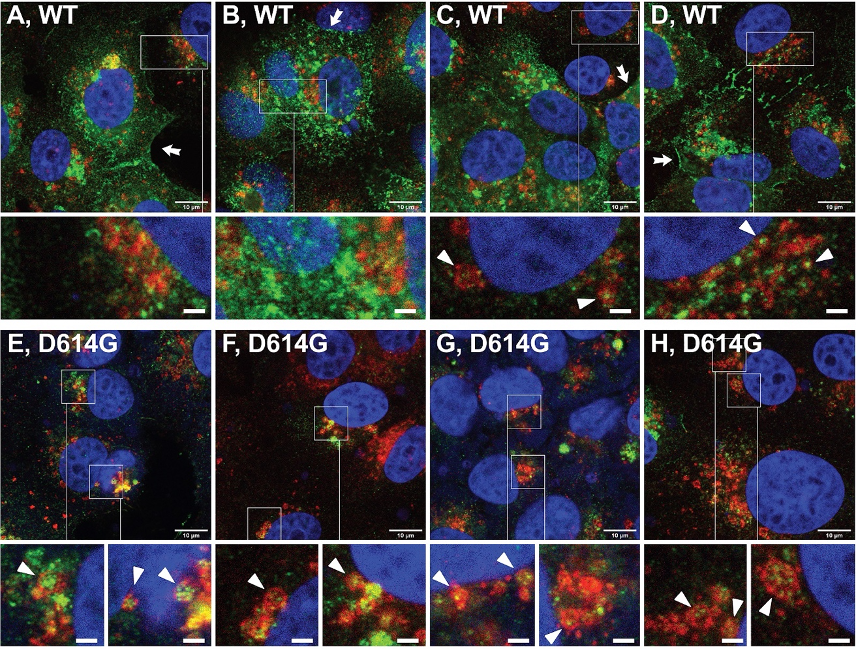
在新冠病毒感染的细胞中,D614G改变S蛋白的运输
至于新冠病毒为什么会发生D614G突变,研究人员认为,该突变出现的时间和重复、独立的起源表明,它的产生正是为了适应新冠病毒进化历程中相对较新的事件,而在新冠病毒的近期历史/史前历史中只发生了两起重大事件:由动物传染到人类,以及新冠病毒特有的FCSI变异。由此推理,D614G出现的原因有两种可能,一种是对其新宿主人类的进化适应,另一种是为了修正先前FCSI所带来的不利影响而产生的进化适应。
研究人员首先在进化史上距离人类较远的哺乳动物负鼠的细胞中表达了S/WT和S/D614G两种蛋白,并测量了S蛋白的总表达量和在细胞表面的表达水平,得出了与之前一致的结论,从而排除了“D614G是人类特有的进化适应”这一可能性。
接下来,Gould团队根据几组不同的实验发现,抑制弗林蛋白酶或破坏FCSI,确实恢复了S蛋白向溶酶体运输的能力,而弗林蛋白酶介导的S蛋白切割,导致S蛋白转运至溶酶体的功能出现明显缺陷,并使细胞表面S蛋白的表达量增加约3倍。研究人员认为,D614G通过还原S蛋白向溶酶体运输的能力,抑制了S蛋白转运中与FCSI相关的有害特性。
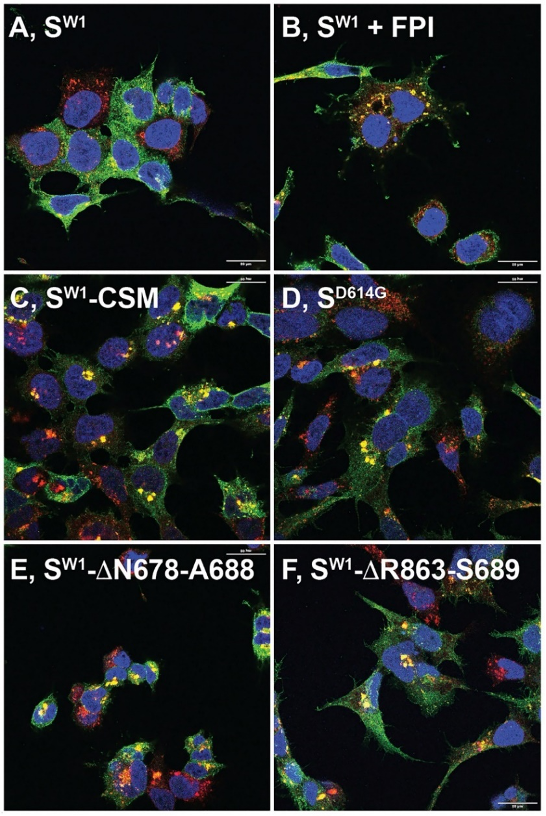
弗林蛋白酶和FCSI破坏了S蛋白向溶酶体的运输
在后续的实验里,研究人员利用遗传学方法发现,S蛋白向溶酶体的转运是由其细胞外结构域调节的。此外,Gould团队基于多个对照试验,提供了首个明确证据表明,仅仅依靠S蛋白的表达就足以驱动溶酶体聚集、KDEL受体在溶酶体积累、ER驻留蛋白的异常分泌,甚至可能阻断内吞物质的溶酶体摄取。
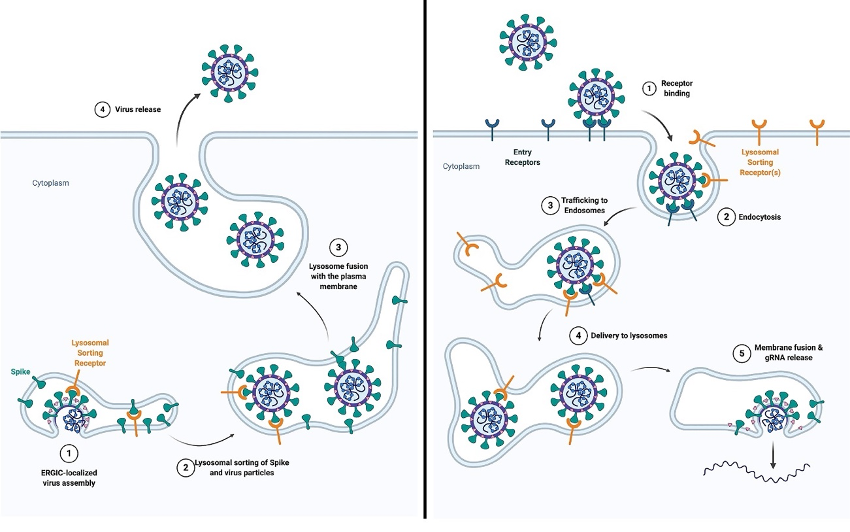
新冠病毒通过胞内体/溶酶体进出细胞的模型
总的来说,从细胞生物学的角度,Gould团队首次证实新冠病毒S蛋白是一种溶酶体蛋白,在β冠状病毒感染的细胞中,除了受体结合、膜融合以外,还能改变溶酶体的结构并重塑溶酶体的功能。
从新冠病毒进化的角度,Gould团队的研究成果表明,最早出现的两个突变,FCSI和D614G,在功能上是密切相关的。D614G突变的出现是为了修复FCSI引起的不良副作用而发生的进化适应,同时依然允许弗林蛋白酶切割S蛋白并支持新冠病毒在呼吸系统传播。

新冠病毒早期进化的遗传模型
Gould团队强调,新冠研究应该拓宽考虑S蛋白突变的遗传和功能背景,虽然所有S蛋白突变都可能代表对其环境的适应,但也有可能是对先前突变的适应。除了研究S蛋白突变对其受体结合、膜融合催化能力的作用,还应该关注这些突变对S蛋白在宿主细胞内的转运及其对S蛋白介导的溶酶体重编程的影响。
参考文献
1.https://www.science.org/doi/10.1126/sciadv.ade5085
2.https://www.biorxiv.org/content/10.1101/2020.12.08.417022v1
3.https://pubmed.ncbi.nlm.nih.gov/32571797/
4.https://pubmed.ncbi.nlm.nih.gov/32568027/
5.https://pubmed.ncbi.nlm.nih.gov/33476327/
6.https://pubmed.ncbi.nlm.nih.gov/33835028/
7.https://pubmed.ncbi.nlm.nih.gov/34159616/
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
