
肝脏再生与类器官形成中表观遗传重塑过程
来源:BioArt 2019-11-14 12:50
在成体肝脏中,生理条件下细胞迭代的速率较低。而肝脏遇到组织损伤的情况下,细胞则能够高效地发挥再生能力【1-4】。最近有研究发现,胆管细胞能够发展成为具有自我更新能力的肝脏类器官,并且具有分化成为肝细胞和导管细胞的能力【5】。但是胆管细胞获得细胞可塑性、起始类器官发育以及应对组织损伤的再生能力是如何发生的,这其中的分子机制还很不清楚。2019年11月4日,剑桥大学Meritx
在成体肝脏中,生理条件下细胞迭代的速率较低。而肝脏遇到组织损伤的情况下,细胞则能够高效地发挥再生能力【1-4】。最近有研究发现,胆管细胞能够发展成为具有自我更新能力的肝脏类器官,并且具有分化成为肝细胞和导管细胞的能力【5】。但是胆管细胞获得细胞可塑性、起始类器官发育以及应对组织损伤的再生能力是如何发生的,这其中的分子机制还很不清楚。
2019年11月4日,剑桥大学Meritxell Huch研究组在Nature Cell Biology发表长文文章Epigenetic remodelling licences adultcholangiocytes for organoid formation and liver regeneration,发现胆管细胞通过全基因组的甲基化/羟甲基化表观遗传景观重塑的分子机制使得分化的细胞可以响应组织损伤从而获得自我更新和分化的能力。
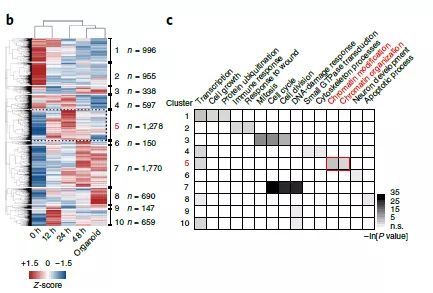
体外培养的胆管细胞类器官具有两种潜能,一种是表达干细胞/祖细胞的标记物,另外一是具有胆管细胞和肝类似细胞的分化能力,能够在移植体内后发育成为成熟的肝细胞【5】。表观遗传机制在发育过程中确保细胞正确分化。在哺乳动物中,DNA的甲基化修饰尤其是5mC (5-methylcytosine)对于基因表达具有重要的作用【6,7】。而DNA的去甲基化可能是由于复制过程中甲基化维持缺失或者是通过TET(Ten-eleven translocation)家族的甲基胞嘧啶加双氧酶活性导致的5mC向5hmC的转化【8】。而5mC的“擦除”以及TET1的活性对于基因组重置从而获得多能性以及体细胞重编程等方面具有非常关键的作用【9】。 作者们从未受损的肝脏中分离出能产生类器官培养的胆管细胞。为了分析胆管细胞起始类器官形成的分子基础,作者们首先对于胆管细胞重新进入细胞周期的时间进行了评估。作者们发现,分离后约40小时后胆管细胞开始进行入S期,约10小时后进入G1期。随后对于刚分离的胆管细胞、分离后进入S期之前以及增殖起始之后的胆管细胞通过RNA-seq进行了全基因组分析。分析后发现,在进入S期之前的最初24个小时,有超过3000个基因的表达发生了变化;而在增殖开始之前,有900个基因的表达发生了变化(图1)。这些结果表明,培养的类器官最初转录水平的变化发生和建立在48小时之内。而对这些发生变化的基因进行聚类后发现,约55%的基因属于表观遗传修饰因子。这给了作者们一个提示,表观遗传机制可能参与到了肝脏类器官起始过程中。
而进行肝损伤处理之后,体内的胆管细胞基因表达也发生了显着地变化,而且细胞增殖明显增加。导管细胞在应对组织损伤的情况时,会对转录组进行一个全基因组的“重写”,因此,该肝脏类器官成为了组织再生分子机制研究的不二之选。 为了进一步确认是哪些表观遗传的因子参与到组织再生过程之中,作者们对于RNA-seq中找到的一些备选基因进行了敲低,发现敲低Tet1之后会显着影响类器官的形成,而敲低Tet2虽然对类器官有影响但该影响并不显着。而且TET1的催化活动对于肝脏类器官的起始和生长是必须的。
考虑到TET1调节羟甲基化的作用,作者们认为DNA的甲基化和羟甲基化的水平可能会参与到体内胆管细胞对于组织损伤的响应。因此,作者们对未损伤以及损伤后的胆管细胞进行了单碱基分辨率的全基因组亚硫酸氢盐测序(Whole-genome bisulphite sequencing ,WGBS)。测序后发现,组织损伤后的胆管细胞中胞嘧啶修饰在全基因组水平均呈现增加的趋势。全基因组的结果表明这种短暂的5hmC的增长可能会是胆管细胞在组织损伤后获得细胞可塑性的原因。 那么这种DNA甲基化修饰的变化是如何调节肝脏细胞再生以及类器官起始的呢?由于TET1在肝脏再生过程中可能参与了细胞可塑性的获得。
因此,作者们使用DamID-seq (DNA adenine methyltransferaseidentification sequencing)技术希望找到TET1在基因组结合的区域,确认TET1通过调节哪些基因的表达促进胆管细胞获得细胞可塑性。同时,作者们还进行了H3K4me3的DamID-seq,发现TET1在肝脏类器官中结合的基因同时也具有H3K4me3的结合峰,说明这些基因大多是转录激活的基因(图2)。而且,TET1通过调节YAP-Hippo、ErbB以及MAPK信号通路促进了胆管细胞可塑性的获得。在小鼠体内的肝脏损伤模型中,也同样发现了TET1会结合在这些基因的转录起始位点,来促进胆管细胞响应组织损伤。
总的来说,Meritxell Huch研究组的工作证明胆管细胞在受到组织损伤以及在随后的类器官形成过程中经历了全基因组DNA甲基化修饰的表观遗传景观的变化。TET1介导的羟甲基化作用对于体内或者体外的胆管细胞激活都具有非常重要的作用,而且胆管细胞可塑性获得过程中也需要YAP-Hippo、ErbB以及MAPK等多种信号通路的参与。该工作为已经分化的细胞应对组织损伤起始细胞再生功能提供了一个可能的、更为广泛的分子机制。(生物谷Bioon.com)
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
