
研究揭示脂肪细胞内质网中错误折叠的蛋白处理新机制
来源:网络 2023-06-20 17:20
本研究利用小鼠模型首次证实了SEL1L-HRD1 ERAD 和ER-phagy在生理条件下的互作。二者共同控制重要生理功能蛋白的折叠与降解,从而维持内质网与细胞稳态。 同时,研究者首次发现脂肪细胞中由
细胞通过各种不同的蛋白质量控制机制(Protein Quality Control Mechanisms)防止有害蛋白聚集。 这些蛋白质量控制机制的退化或故障导致众多人类疾病,如糖尿病、癌症,和各种老年疾病等。内质网作为蛋白质折叠和质量控制的主要场所,利用内质网蛋白降解 (ERAD) 和内质网自噬 (ER-phagy)两种机制降解错误蛋白。其中ERAD 有针对性地降解内质网中的蛋白,而ER-phagy,作为自噬 (autophagy) 的一种,常被认为批量降解已碎片化的内质网或内质网域 (ER fragments/domains)。 然而,作为两种最重要的蛋白降解机制, 我们对ERAD和ER-phagy的作用机制,尤其是在生理条件下二者如何协同并维护内质网稳态仍知之甚少。
2023年5月31日, 密歇根大学医学院齐岭教授课题组及清华大学生命科学院,清华-IDG/麦戈文脑科学研究院吝易助理教授课题组在《自然通讯》(Nature Communications)杂志上在线发表题为“脂肪细胞中内质网处理错误折叠蛋白机制”( The mechanisms to dispose of misfolded proteins in the endoplasmic reticulum of adipocytes )的研究论文。研究揭示了在小鼠脂肪细胞中SEL1L-HRD1 ERAD和ER-phagy的共同作用机制,并发现脂肪细胞对错误折叠蛋白在内质网中的积累可能具有高度适应性。正常状态下ER-phagy功能受到限制,SEL1L-HRD1 ERAD为降解错误折叠蛋白核心机制。 当SEL1L-HRD1 ERAD功能下降时,脂肪细胞中ER-phagy功能增加,从而减少错误折叠蛋白形成的聚集物在内质网中积累。而当SEL1L-HRD1 ERAD 和ER-phagy 功能均下降的情况下,错误折叠的蛋白可以被储存在一种由内质网碎片聚集融合形成的特殊细胞结构中。这种细胞结构被命名为Coalescence of ER Fragments (CERFs)。 研究者推测此种结构可能对脂肪细胞 (至少在短期内)适应蛋白质量控制机制缺陷具有重要作用。
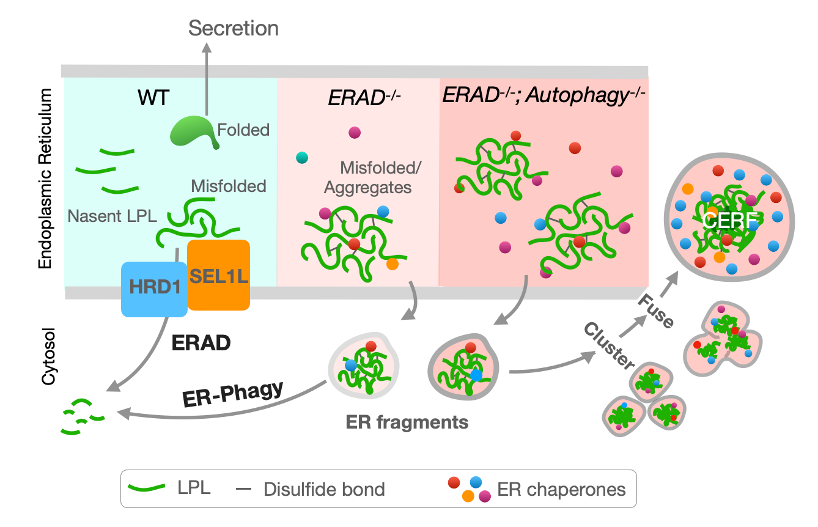
图1 – 小鼠脂肪细胞有步骤地处理内质网中错误折叠蛋白及蛋白高聚物的假设模型。

图2 – 透射电子显微镜(TEM)下观察到在脂肪细胞特异SEL1L-HRD ERAD 和ER-phagy 双敲除(DKO)的小鼠的脂肪组织里形成的CERFs结构 (a, 箭头:在CERFs中的类膜结构)。使用 免疫金染色后,在TEM下观察到CERFs结构 (绿色虚线) 富集着内质网腔标记蛋白BiP (b)。 LD: lipid droplet.
随后,研究者利用免疫沉淀质谱法(immunoprecipitation-mass spectrometry)分析了小鼠脂肪组织中CERFs的主要成分。结合一系列生化分析,他们发现脂蛋白脂肪水解酶- lipoprotein lipase (LPL)是CERFs主要的蛋白成分,也是脂肪细胞中重要的ERAD底物。
LPL 主要由脂肪与肌肉细胞分泌,其主要功能是水解在乳糜微粒(Chylomicrons)和极低密度脂蛋白(VLDL)里的甘油三酯。LPL 的功能缺失在严重的情况下可以导致I型高脂蛋白血症,而较轻程度上则引起体内脂蛋白代谢的紊乱。因此,阐明LPL蛋白在细胞内的合成机制将为了解、预防或治疗高血脂疾病做出重要贡献。
研究发现,LPL在内质网中的折叠主要受SEL1L-HRD1 ERAD控制。ERAD可及时清除在内质网中错误折叠的LPL蛋白,从而确保正确折叠的LPL蛋白被正常分泌。在ERAD功能缺失情况下,LPL会形成蛋白高聚物;然而,此时脂肪细胞可激活ER-phagy作为ERAD 缺失的补偿机制,限制LPL在内质网中聚集。若脂肪细胞中ERAD和ER-phagy功能双重缺失,错误折叠LPL 蛋白将被储存在CERFs, 并随着时间形成低溶解度的高聚物。
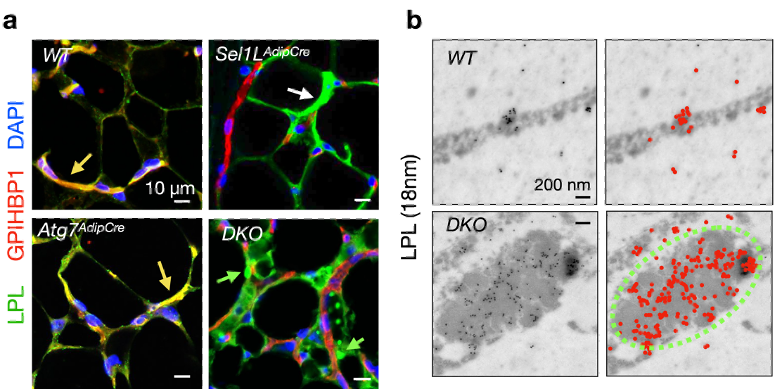
图3 – 使用免疫荧光实验在共聚焦显微镜下观察到,在野生型(WT)和自噬单敲(Atg7AdipCre)的小鼠中,LPL可被脂肪细胞分泌并进入血管中,从而与血管标记蛋白GPIHBP1共定位(a,黄色箭头); 然而在SEL1L-HRD1 ERAD单敲(Sel1LAdipCre)和ERAD与自噬双敲(DKO)的小鼠中, LPL累积在脂肪细胞中从而不能被正常分泌到血管中(a, 白色箭头)。相比于SEL1L-HRD1 ERAD单敲(Sel1LAdipCre),在双敲小鼠(DKO)中,LPL不仅在脂肪细胞中累积,大部分且形成圆形焦点(a, 绿色箭头)。使用免疫金染色特异标记LPL后,可在TEM下观察到LPL在双敲脂肪细胞中富集于CERFs结构 (绿色虚线)中(b)。
研究还发现了CERFs中蛋白的特异分布形态。荧光和免疫金电镜成像结果均显示在CERFs中,LPL被内质网中的分子伴侣BiP包裹。为了验证CERFs组装的机制,研究者对CERFs进行了体外重构。结果显示,纯化的 LPL蛋白具有自聚集的能力,能够在体外自发相分离并形成中空的具有类似胶状性质的液滴。另外, 纯化的BiP可与LPL一起相分离并在体外形成BiP包裹LPL的液滴。这些发现将为未来进一步阐明LPL在细胞内的聚集和调控机制打下基础。
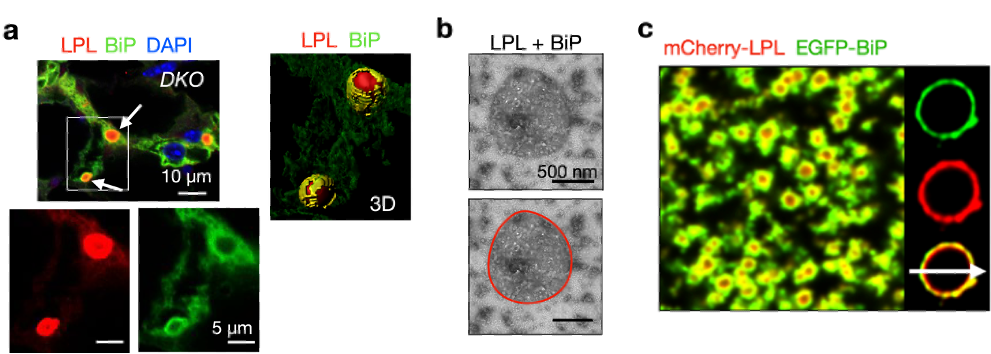
图4 – 使用免疫荧光实验在共聚焦显微镜下观察到,在ERAD与自噬双敲的小鼠(DKO)脂肪组织中,LPL CERFs的外围被BiP 环绕 (a,箭头)。负染色后,在TEM下观察到纯化BiP与LPL一起相分离(b)。荧光染色后,发现纯化的BiP包裹LPL液滴(c)。
综上,本研究利用小鼠模型首次证实了SEL1L-HRD1 ERAD 和ER-phagy在生理条件下的互作。二者共同控制重要生理功能蛋白的折叠与降解,从而维持内质网与细胞稳态。 同时,研究者首次发现脂肪细胞中由内质网碎片聚集融合形成的特殊细胞结构CERFs, 揭示了其生理功能,并解析了CERFs的主要蛋白成分及组装机制。未来研究将进一步探究 ERAD,ER-phagy 和CERFs形成在由蛋白折叠缺陷引起的疾病中的重要性。
密歇根大学医学院齐岭教授, 清华大学生命科学院,清华-IDG/麦戈文脑科学研究院吝易教授为本文共同通讯作者。密歇根大学博士生吴霜程(Alivia)为第一作者。清华大学生命科学院申晨晨进行了体外试验部分。其他密歇根大学医学院齐岭实验室科研同事协助实验。
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
