
Nature子刊:理性设计构建合成能量系统双引擎助力细胞工厂
来源:深圳先进院 2022-11-03 11:06
生物技术的挑战之一是如何在刚性代谢网络的基础上改变胞内能量(ATP)和还原力(NADH/NADPH)的相对化学计量比,来积累过量能量/还原力进行产品合成。
于涛团队致力于利用合成生物学方法,解决可持续制造、绿色能源的生物存储与粮食安全等重大问题。前期工作中,团队成功实现了空气中CO2到葡萄糖与脂肪酸的制备(Nature Catalysis,2022),该工作开发的合成能量系统(细胞双引擎),进一步提高了脂肪酸的转化率,并为下一步开发可再生能源的生物储能技术打下了基础。
细胞在生长过程中合成大分子、构建碳骨架,这些过程消耗能量,且需要克服底物和生物量之间的还原度差异。对于脂质等高价值的储能化合物,其还原度远高于葡萄糖等底物,细胞往往需要额外的还原力和能量来合成这些化合物。而这无疑需要对细胞的代谢网络进行重构,因为细胞获取能量和还原力、合成大分子和构建碳骨架的目的是为自身生长,而不是产品合成。因此,研究人员构想,是否有可能从头构建一个合成的能量系统,其是否可以取代内源的能量系统为细胞供应能量支撑生长?其是否能为化学品合成提供额外的能量和还原力?细胞内的反应高度协作,胞内的能量和物质组成维持在一定的比例,细胞是否能接受过量的NADPH和NADH?
研究团队通过理性重排还原代谢,在酵母细胞内构建了一个新型合成能量系统,利用重复的单脱羧反应和转氢循环实现NADPH和NADH的合成,并进一步转变为能量进行释放。该系统由三个模块组成:磷酸戊糖(Pentose Phosphate,PP)循环、转氢循环和外部呼吸链(图1)。
重复脱羧是细胞还原代谢的基础,因此要构建一个新的能量系统首先就是要构建一个重复脱羧循环。磷酸戊糖途径中,6-磷酸葡萄糖(G6P)被催化为5-磷酸核糖(R5P)时失去一分子CO2,生成2分子NADPH,此为氧化阶段;之后C5分子经可逆重排,最终重构为6-磷酸果糖(F6P)和3-磷酸甘油醛(G3P),此为非氧化阶段;G3P经部分糖异生途径可以回到G6P,再一次进入氧化阶段,这样就构成了PP循环。在这个循环中,1分子葡萄糖彻底氧化可以生成12分子NADPH。NADPH是细胞内的还原力通量,而NADH经过氧化呼吸链可以生成ATP。研究人员过表达酵母内源谷氨酸盐转氢酶GDH1和GDH2,在胞质内实现1分子NADPH不可逆转变为1分子NADH的转氢循环。
研究人员选择了一株消除葡萄糖效应的丙酮酸脱羧酶缺陷菌株E1B[1](重命名为SynENG001)来测试PP循环和转氢循环的碳通量。该菌株以氧化磷酸化为主要供能过程,可以在单一葡萄糖碳源上生长,且具有较好的胞质NADH平衡能力。研究人员敲除了磷酸果糖异构酶基因pgi1,打断了糖酵解途径,提高了PP循环碳通量,得到的菌株SynENG064不能以糖为单一碳源生长;而在添加转氢循环后,得到的菌株SynENG065在5天内可以生长到OD600 nm=10(图2b)。由此可推测敲除pgi后胞内的NADPH过量,对细胞生长产生抑制。在野生型酵母内敲除pgi并过表达GDH2仅能部分恢复细胞生长,研究人员推测这是由于转氢循环产生的过量NADH抑制细胞生长,因此细胞需要一个有效的呼吸链来解除胞质内多余的NADH。以上结果证实PP循环和转氢循环运行良好,且两个模块有较高的碳通量。
琥珀酸是一种四碳二羧酸,从丙酮酸合成琥珀酸需要细胞提供额外的还原力NADH。研究人员在SynENG001菌株中过表达了琥珀酸合成所需要的各基因,并下调了磷酸果糖激酶(PFK)的表达水平推动碳流更多流向PP循环,使琥珀酸的产量升至约3.3 g/L。在此过程中研究人员还发现了甘油的积累,这些都说明PP循环和转氢循环导致细胞内的NADH过剩。
研究人员模拟线粒体内的氧化呼吸链,表达了两种外部NADH脱氢酶NDE1和NDE2,将胞质NADH与线粒体电子传递链联系起来,得到菌株SynENG012。在下调TCA循环关键基因异柠檬酸脱氢酶IDH2的表达水平时,细胞生长逐渐受损乃至致死;放入合成能量系统(转氢循环和外部呼吸链)后,细胞生长不受影响。这些说明合成能量系统可以替代胞内原有能量系统发挥作用,支持细胞生长。
自然界的产油真菌中线粒体内的异柠檬酸脱氢酶受损会导致油脂的过度生产,因此先前的研究中,研究人员在已优化的产自由脂肪酸(Free Fatty Acids,FFAs)菌株动态调控IDH2的表达,得到菌株Y&Z032[2]。但是摇瓶发酵后细胞生长和FFAs产量都下降了。研究人员推测这是由于胞内能量供应不足,因此放入了转氢循环和外部呼吸链,得到的菌株SynENG024生物量相对提高100%,FFA产量提高200%。这也进一步印证了合成能量系统可以支持细胞生长和高还原性化合物的生产。
此外,研究人员还过表达不同来源的果糖-1,6-二磷酸酶FBP1来提高NADPH供应;使用不同的启动子微调NADPH和ATP的比例,但并未观察到FFAs产量的明显提高。因此在胞内表达了非氧化糖酵解(Non-oxidative glycolysis,NOG)途径,将PP途径部分分流到乙酰辅酶A的合成。如预期所料,FFAs产量提高了30%。这说明最优生产需要结合多策略实现。
分批补料发酵实验中,研究人员在FFAs生产的最佳菌株SynENG050中恢复了完整功能的TCA循环来避免高浓度乙醇的积累,又过表达一份FFAs合成途径来竞争碳源和消耗过多的NADH,得到菌株SynENG058,发酵后使用Dodecane进行提取,得到20 g/L的FFAs,产率达到0.134 g FFAs/g葡萄糖,为最高理论得率的40%。
生物技术的挑战之一是如何在刚性代谢网络的基础上改变胞内能量(ATP)和还原力(NADH/NADPH)的相对化学计量比,来积累过量能量/还原力进行产品合成。该研究构建了一个合成能量系统,为细胞供给额外的能量和还原力,经过实验验证,该系统可替代原有的TCA循环,可支持高还原度化学品的生产,也可与其他途径合作高效生产以乙酰辅酶A为前体的化学品。这是第一个通过理性设计构建的合成能量系统。这项研究揭示了细胞能量代谢网络的可塑性:尽管经过了几百万年的进化,细胞的能量代谢网络依然可以重构。
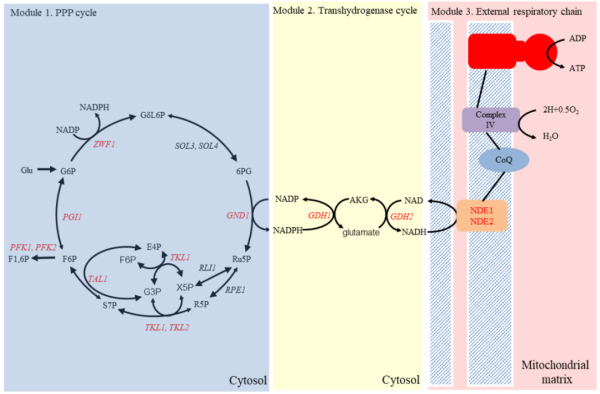
图1 合成能量系统示意图
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
