
研究揭示病毒RdRP核苷酸催化循环机制
来源:武汉病毒所 2020-05-26 20:30
RNA病毒是一类独特的生命形式,其基因组复制过程完全不涉及DNA形式,因此需要自身编码的依赖RNA的RNA聚合酶(RNA-dependent RNA polymerase,简称RdRP)来主导完成。对多数RNA病毒来说,基因组复制过程缺乏纠错机制,具有较高的变异性,因此增加了疫苗和药物研究的整体难度。深入研究RdRP的催化与调控机制,对了解RNA
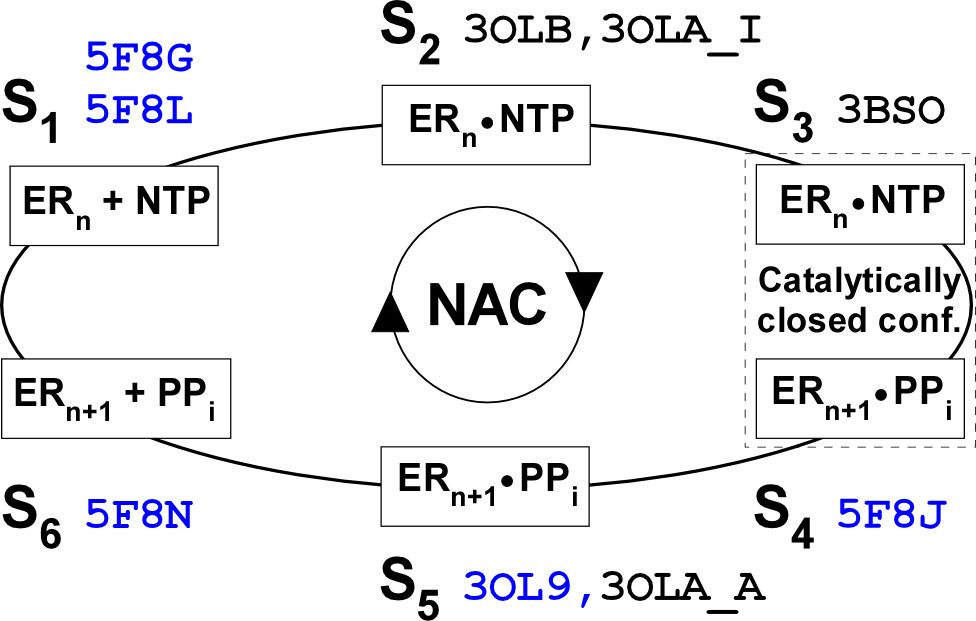
RNA病毒是一类独特的生命形式,其基因组复制过程完全不涉及DNA形式,因此需要自身编码的依赖RNA的RNA聚合酶(RNA-dependent RNA polymerase,简称RdRP)来主导完成。对多数RNA病毒来说,基因组复制过程缺乏纠错机制,具有较高的变异性,因此增加了疫苗和药物研究的整体难度。深入研究RdRP的催化与调控机制,对了解RNA病毒的本质、掌握其进化规律和发展病毒防控策略至关重要。RdRP催化过程主要由链引发和链延伸两个阶段构成。其中不稳定的引发阶段主要实现对合成起始位点的精确控制,进而在合成数个核苷酸后完成向稳定的延伸阶段的转换,延伸阶段由数以千计且机制相同的核苷酸添加循环构成,因此核苷酸添加循环机制是RdRP催化RNA合成的核心内容。
RdRP最早于上世纪六十年代早期在小RNA病毒科的门戈病毒和脊髓灰质炎病毒等的相关研究中被发现(Reich et al. Science 1961;Reich et al. PNAS 1962)。从1997年Steve Schultz研究组报道脊髓灰质炎病毒RdRP晶体结构(Hansen et al. Structure 1997)至今,已有数十种病毒的RdRP三维结构获得解析。与其他类型的单亚基聚合酶相似,RdRP的核心区形似杯装右手,由手掌(palm)、拇指(thumb)和手指(fingers)三个结构域围绕形成一个催化中心。与其他单亚基聚合酶不同的是,RdRP拇指与手指尖端通过疏水相互作用形成了环绕式(encircled)右手结构。后续研究表明,RdRP这一独特环绕式结构很可能与其催化机制独特性相关,由于手指区的运动性受到了拇指与手指相互作用的限制,病毒RdRP在底物NTP诱导的催化中心关闭过程中的构象变化主要发生在手掌结构域(Gong and Peersen PNAS 2010),而其他类型的单亚基聚合酶的相关构象变化则主要发生在手指区(Huang et al. Science 1998; Li et al. EMBO J 1998; Yin and Steitz Cell 2004)。
中国科学院武汉病毒研究所研究员龚鹏长期从事病毒RdRP的催化与调控机制研究,前期在Olve Peersen研究组从事博士后研究期间揭示了RdRP基于手掌区构象变化的催化中心关闭特征,提出了六状态核苷酸添加循环模型和转位中间体假说(图1)。2011年成立研究组以来,继续致力于阐明RdRP核苷酸添加循环机制,2016年利用时间分辨晶体学等方法进一步揭示了催化中心关闭由两个步骤构成,其中第一步构象变化由NTP核糖2¢位羟基诱导,从而阐明了2¢位修饰的核苷类分子(如丙型肝炎特效药索非布韦)可特异性抑制RNA病毒的结构基础(Shu and Gong PNAS 2016)。在此项工作中,该团队还捕获了一种转位中间体(translocation intermediate)结构,发现RdRP在完成催化反应后RNA产物链先于模板链向上游移动,而RdRP独特的催化基序G(motif G)与模板链的“锁定”相互作用很可能是造成RNA双螺旋不对称运动的主要因素。
为充分揭示RdRP转位机制和完整绘制RdRP核苷酸添加循环图,该研究团队充分结合晶体学与酶学方法开展研究,通过改变肠道病毒RdRP-RNA复合物晶体浸泡实验中的底物NTP和副产物焦磷酸的浓度,同时结合晶体浸泡时间扫描策略,获得了正向转位和逆向转位的中间体结构。在这两类中间体结构中,RNA产物链均先于模板链发生移动,进一步提示了基序G的“锁定”作用对转位过程的控制(图2,PDB号:6LSE、6LSF)。与正向转位中RNA双链始终维持原有的碱基对相互作用不同,逆向转位中间体提示RNA双链见发生“滑动”,导致原有碱基对相互作用被打破,重新建立了亚稳定的不完全配对的双链。这一结果一定程度上提示了聚合酶催化的逆向反应比正向反应更难发生,同时也为理解聚合酶的纠错功能提供了重要依据。
为了观测到转位末期基序G“锁定”作用如何打破,从而允许模板链发生移动,研究团队系统性分析了基序G参与锁定的两个关键氨基酸位点(对应肠道病毒RdRP的114和115位),发现大多数RdRP在这两个位点均使用四种小侧链氨基酸(G:甘氨酸;A:丙氨酸;S:丝氨酸;T:苏氨酸),推测上述“锁定”作用很可能由空间位阻实现控制。在测试了这两个位点的G/A/S/T四种氨基酸的全部16种组合后,发现多数突变体RdRP仍然可以维持催化功能,但催化效率(由kcat/KM,NTP评测)比之野生型发生不同程度的变化,提示突变可能对“锁定”作用产生调节(图3)。研究团队通过进一步尝试突变体RdRP-RNA复合物结构解析,利用一种kcat和野生型相当但KM,NTP五十倍于野生型(即NTP亲和力远低于野生型)的突变体成功获得了模板链“解锁”的转位后期中间体结构(图4,PDB号:6LSG、6LSH)。
至此,龚鹏研究组系统性阐释了RdRP核苷酸添加循环中化学反应前的催化中心关闭和化学反应后的转位这两个重要构象变化的精细过程,相对完整地绘制了循环图。为进一步研究RdRP错配、基因组复制纠错和研制针对RNA病毒的核苷类高效和特效药物奠定了重要基础。研究团队进一步提出,RdRP通过“锁定”相互作用对转位实行严格控制很可能是为了保障模板碱基的精确定位和高效催化,类似的模板锁定以及转位与催化的偶联机制很可能在其他类型的聚合酶中同样存在。(生物谷Bioon.com)
版权声明 本网站所有注明“来源:生物谷”或“来源:bioon”的文字、图片和音视频资料,版权均属于生物谷网站所有。非经授权,任何媒体、网站或个人不得转载,否则将追究法律责任。取得书面授权转载时,须注明“来源:生物谷”。其它来源的文章系转载文章,本网所有转载文章系出于传递更多信息之目的,转载内容不代表本站立场。不希望被转载的媒体或个人可与我们联系,我们将立即进行删除处理。
